figura 4.1 (ver vista ampliada)
En consecuencia, una comprensión de esta sinapsis conduce a una comprensión de las otras. Por lo tanto, primero discutiremos el proceso de transmisión sináptica en la unión neuromuscular esquelética.,
Las características de la Unión sináptica en la unión neuromuscular se muestran en la figura de la izquierda. Las fibras del músculo esquelético son inervadas por neuronas motoras cuyos cuerpos celulares están ubicados en el cuerno ventral de la médula espinal. La región terminal del axón da lugar a procesos muy finos que se ejecutan a lo largo de las células del músculo esquelético. A lo largo de estos procesos se encuentran estructuras especializadas conocidas como sinapsis. La sinapsis particular hecha entre una neurona motora espinal y la célula del músculo esquelético se llama la placa final motora debido a su estructura específica.,
la sinapsis en la unión neuromuscular tiene tres características características de las sinapsis químicas en el sistema nervioso. En primer lugar, hay una clara separación entre la membrana presináptica y la postsináptica. El espacio entre los dos se conoce como hendidura sináptica. El espacio nos dice que debe haber algún mecanismo de señalización intermedio entre la neurona presináptica y la neurona postsináptica para que la información fluya a través de la hendidura sináptica. En segundo lugar, hay una alta densidad característica de pequeñas vesículas esféricas., Estas vesículas sinápticas contienen sustancias neurotransmisoras. Las sinapsis también están asociadas con una alta densidad de mitocondrias. En tercer lugar, en la mayoría de los casos, hay un engrosamiento característico de la membrana postsináptica, que se debe al menos en parte al hecho de que la membrana postsináptica tiene una alta densidad de receptores especializados que se unen a las sustancias químicas transmisoras liberadas de la neurona presináptica. En los capítulos 8 y 10 se proporcionan detalles adicionales sobre las características morfológicas de las uniones sinápticas.
4.,2 fisiología de la transmisión sináptica en la unión Neuromuscular
figura 4.2
la figura de la derecha ilustra de manera muy esquemática cómo es posible estudiar con gran detalle la fisiología de la transmisión sináptica en la unión neuromuscular esquelética. Un trozo de músculo y su nervio conectado se colocan en una pequeña cámara experimental llena de una solución de timbre adecuada. El potencial de reposo de la célula muscular se registra con un microelectrodo. También se colocan electrodos en la superficie del axón nervioso., Las descargas eléctricas breves hacen que se inicien potenciales de acción, que se propagan al terminal sináptico.
la figura siguiente ilustra dos tipos de cambios potenciales que se registraron en una preparación nervio-músculo aislada. El experimento también ilustra las propiedades de un poderoso fármaco, el curare, que ha demostrado ser muy útil en el estudio del proceso de transmisión sináptica en la unión neuromuscular esquelética. La parte a ilustra la secuencia de cambios potenciales registrados en la célula muscular como resultado de la estimulación del axón motor., La flecha indica el punto en el tiempo cuando el choque se entrega al axón motor. Tenga en cuenta que hay un período de tiempo inactivo después del choque. El retraso se debe al tiempo que tarda el potencial de acción en el axón motor en propagarse desde su sitio de iniciación. Después del retraso, hay dos tipos de potenciales registrados en la célula muscular. En primer lugar, hay un potencial de cambio relativamente lento que será el Centro de la siguiente discusión., Si ese potencial inicial lento es suficientemente grande, como lo es normalmente en las células del músculo esquelético, un segundo potencial, un potencial de acción, se produce en la célula muscular.
figura 4.3
los potenciales de acción en las células del músculo esquelético se deben a mecanismos iónicos similares a los discutidos anteriormente. Específicamente, hay un cambio dependiente del voltaje en la permeabilidad de Na+ seguido de un aumento retardado en la permeabilidad de K+. (Para las células del músculo liso y las células del músculo cardíaco los mecanismos iónicos son diferentes, sin embargo.,)
el evento subyacente que desencadena el potencial de acción puede revelarse aprovechando el curare, un veneno de flecha utilizado por algunos indios sudamericanos. Una dosis baja de curare (Parte B) reduce el evento subyacente, pero todavía no se reduce lo suficiente para caer por debajo del umbral. Si se administra una dosis algo más alta de curare (Parte C), el evento subyacente lento se convierte en subtreshold. La señal subyacente se conoce como el potencial de la placa final (EPP) porque es un cambio de potencial registrado en la placa final del motor. Generalmente, se conoce como potencial postsináptico excitatorio (EPSP).,
Curare bloquea el potencial de la placa final porque es un inhibidor competitivo de la acetilcolina (ACh), el transmisor liberado en el terminal presináptico. Curare no bloquea la conductancia dependiente de voltaje Na+ o la conductancia dependiente de voltaje K+ que subyace al potencial de acción muscular. Curare afecta el estímulo (el EPSP) que normalmente conduce a la iniciación del potencial de acción muscular. Un animal envenenado con curare se asfixiará porque el proceso de transmisión neuromuscular en los músculos respiratorios está bloqueado.,
normalmente, la magnitud del potencial endplate es bastante grande. De hecho, la amplitud del potencial de la placa final es de aproximadamente 50 mV, pero solo se necesitan alrededor de 30 mV para alcanzar el umbral. Los 20 mV adicionales se denominan factor de seguridad. Por lo tanto, incluso si el potencial de la placa final se volviera algo más pequeño (por ejemplo, 40 mV en amplitud) debido a la fatiga, el EPP alcanzaría el umbral y la relación uno a uno entre un potencial de acción en el axón motor y un potencial de acción en la célula muscular se preservaría.
la Figura 4.4
4.,3 propagación del EPP
¿Cuáles son las propiedades del EPP y cómo se compara con las propiedades del potencial de acción?
¿el potencial de la placa final se debe a un cambio dependiente del voltaje en las permeabilidades Na+ y K+ como el potencial de acción?
¿se propaga el potencial endplate de una manera de todo o nada como el potencial de acción?
la figura de la izquierda ilustra un experimento que examina la propagación del potencial endplate. La fibra muscular se empala repetidamente con electrodos a intervalos de 1 mm., (Tenga en cuenta que el potencial de la placa final es pequeño porque este experimento se realiza en presencia de una baja concentración de curare por lo que el potencial de la placa final se puede registrar sin las complicaciones de desencadenar un potencial de acción.) El potencial de la placa final no se propaga de una manera de todo o nada. Se extiende a lo largo del músculo, pero lo hace con decremento., Por lo tanto, la propagación del potencial endplate desde su sitio de iniciación a otros sitios a lo largo de la célula muscular ocurre pasivamente y con decremento, al igual que un cambio de potencial Sub-umbral en una porción del axón se extiende a lo largo del axón, o simplemente como un cambio en la temperatura en un punto en una varilla metálica se extiende a lo largo de la varilla.
4.4 Descripción general de la secuencia de eventos subyacentes al EPP
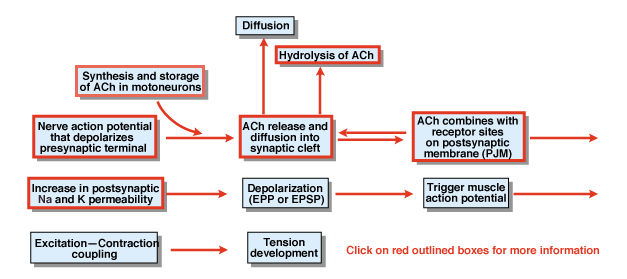
figura 4.5
¿Cuáles son los otros pasos en el proceso de transmisión química sináptica? Gráfico 4,5 proporciona una visión general. Un potencial de acción nerviosa que se inicia en el cuerpo celular de una neurona motora espinal se propaga a las raíces ventrales y eventualmente invade las terminales sinápticas de las neuronas motoras. Como resultado del potencial de acción, el transmisor químico acetilcolina (ACh) se libera en la hendidura sináptica. La ACh se difunde a través de la hendidura sináptica y se une a receptores especiales en la postsináptica o la membrana postsináptico. La Unión de ACh a sus receptores produce un cambio conformacional en un canal de membrana que es específicamente permeable tanto a Na+ como A K+., Como resultado de un aumento en la permeabilidad de Na+ y K+, hay una despolarización de la membrana postsináptica. Esa despolarización se denomina potencial endplate o, más generalmente, EPSP. Si la EPSP es lo suficientemente grande, como lo es normalmente en la unión neuromuscular, conduce a la iniciación de un potencial de acción en la célula muscular. El potencial de acción inicia el proceso de acoplamiento de contracción de excitación y el desarrollo de tensión. La duración del potencial de la placa final es de aproximadamente 10 mseg.
dos factores controlan la duración de la EPSP en la unión neuromuscular., En primer lugar, ACh se elimina por difusión. En segundo lugar, una sustancia en la hendidura sináptica, llamada acetilcolinesterasa (AChE), hidroliza o descompone la ACh. La AChE es una de las enzimas más eficientes conocidas. Una sola molécula de AChE puede hidrolizar 600.000 moléculas de ACh por minuto.
4.5 Papel de la ace
la Figura 4.6
Una importante familia de sustancias, una de las cuales es la neostigmina, inhibe la acción de la AChE. La neostigmina bloquea la acción del dolor y, por lo tanto, hace que el potencial de la placa final sea más grande y de mayor duración., Esta figura ilustra dos potenciales endplate. Una se registró en solución salina y curare y una segunda después de la adición de neostigmina a la solución. (Se añade Curare para que las propiedades del EPP puedan estudiarse sin desencadenar un potencial de acción en la célula muscular.) Después de aplicar la neostigmina, el potencial de la placa final es mucho mayor y de mayor duración.
4.6 A miastenia Gravis
la miastenia gravis se asocia con debilidad muscular severa debido a una disminución en el número de receptores de acetilcolina en la célula muscular., Si la tapa potencial es menor, la tapa potencial de no alcanzar el umbral. Si no alcanza el umbral, no habrá potencial de acción en la célula muscular y no habrá contracción del músculo, lo que causa debilidad muscular. La neostigmina y otros inhibidores de la AChE se utilizan para tratar a pacientes con miastenia grave. Estos agentes hacen que la cantidad de acetilcolina que se libera con mayor eficacia llegar a los restantes receptores de acetilcolina.
4.,6b agentes nerviosos
aunque los inhibidores de la AChE tienen un valor terapéutico importante, algunos inhibidores han sido y siguen siendo utilizados como venenos. Algunos inhibidores de la AChE como el Somán y el sarín forman un bloqueo bastante irreversible de la AChE. Este bloqueo conduce a niveles extremos de ACh en la hendidura sináptica. Las personas así envenenadas mueren de convulsiones y espasticidad muscular, incluidos los músculos respiratorios.
la Figura 4.7
4.,7 iontoforesis de ACh
la iontoforesis es una técnica interesante que se puede utilizar para probar más a fondo la hipótesis de que ACh es la sustancia neurotransmisora en la unión neuromuscular. Si ACh es el transmisor que es liberado por esta sinapsis, uno podría predecir que debería ser posible sustituir la aplicación artificial del transmisor por la liberación normal del transmisor. Dado que la ACh es una molécula con carga positiva, puede ser forzada a salir de un microelectrodo para simular la liberación de ACh desde un terminal presináptico.
Figura 4.,8
de hecho, pequeñas cantidades de ACh se pueden aplicar a la vecindad de la unión neuromuscular. La figura 4.8 compara una EPP producida por la estimulación del axón motor y la respuesta a las eyecciones de ACh. El cambio de potencial parece casi idéntico al potencial de la placa final producido por la liberación normal de ACh. Este experimento proporciona soporte experimental para el concepto de que ACh es el transmisor natural en esta sinapsis.,
la respuesta a la eyección de ACh tiene algunas otras propiedades interesantes que son todas consistentes con la naturaleza colinérgica de la sinapsis en la unión neuromuscular esquelética. La neostigmina hace que la respuesta a la iontoforesis de ACh sea más larga y más grande. Curare reduce la respuesta porque compite con la Unión normal de ACh. Si la ACh es expulsada hacia la célula muscular, no sucede nada porque los receptores de acetilcolina no están en el interior; están en el exterior de la célula muscular., La aplicación de acetilcolina a regiones del músculo alejadas de la placa terminal no produce respuesta porque los receptores de la ACh se concentran en la región sináptica.
para probar su comprensión hasta ahora, considere cómo un agente como TTX afectaría la generación de un EPP y la respuesta de una fibra muscular a la aplicación iontoforética de ACh? TTX no tiene ningún efecto en la respuesta a ACh, pero bloquea el EPP. La razón por la que la respuesta a ACh no se ve afectada es clara, pero muchos esperan que si no hay efecto aquí, tampoco debería haber efecto en el PPE., La tetrodotoxina no afecta la Unión de la acetilcolina a los receptores y por lo tanto no afectará la respuesta a la aplicación directa de ACh. Sin embargo, la tetrodotoxina afectará la capacidad de un potencial de acción para ser provocada en el axón motor. Si no se puede obtener un potencial de acción en el axón motor, no puede causar la liberación del transmisor. Por lo tanto, la tetrodotoxina aboliría totalmente el PPE. El bloqueo no sería debido a un bloqueo de receptores ACh, sino más bien a un bloqueo de algún paso antes de la liberación del transmisor.
4.,8 mecanismos iónicos del EPP
Bernard Katz y sus colegas fueron pioneros en investigar los mecanismos de transmisión sináptica en la unión neuromuscular. Sugirieron que el canal abierto por ACh era uno que tenía igual permeabilidad tanto a Na+ como A K+. Debido a que era igualmente permeable a Na+ y K+, Katz sugirió que, como resultado de la apertura de estos canales, el potencial de membrana se movería hacia 0 mV. (Un valor de alfa en la ecuación de GHK igual a uno, que cuando se sustituye en la ecuación, produce un potencial de aproximadamente 0 mV.,)
la Figura 4.9
El experimento que se muestra en la figura de la izquierda pruebas de concepto. La célula muscular ha sido penetrada con un electrodo de registro, así como otro electrodo que se puede conectar a una fuente adecuada de potencial para cambiar artificialmente el potencial de membrana. Normalmente, el potencial de membrana es de aproximadamente -80 mV nuevamente, se agrega una pequeña cantidad de curare para que el EPP sea pequeño. Katz notó en estos experimentos que el tamaño del EPP cambió dramáticamente dependiendo del potencial de la célula muscular., Si el potencial de membrana se mueve a 0 mV, no se registra ningún cambio de potencial. Si el potencial de membrana se hace +30 mV, el EPP se invierte. Así que tres estímulos diferentes producen potenciales de placa final que son muy diferentes entre sí.
la falta de respuesta cuando el potencial está a 0 mV es particularmente informativa. Considere por qué no se registra ningún cambio potencial. Presumiblemente, el transmisor está siendo liberado y unido a los receptores., La explicación simple de la falta de cambio potencial es que el potencial al que la apertura de los canales ACh están tratando de alcanzar ya se ha logrado. Si el potencial de membrana se hace más positivo que 0 mV, entonces el EPP se invierte. No importa cuál sea el potencial, el cambio en la permeabilidad tiende a mover el potencial de membrana hacia 0 mV! Si el potencial de reposo es más negativo que 0 mV, hay una desviación hacia arriba. Si es más positivo, hay una desviación hacia abajo. Si ya está a 0 mV, no hay desviación.
Figura 4.,10
Este potencial también se llama potencial de reversión, porque es el potencial en el que se invierte el signo del potencial sináptico. El experimento indica que, como resultado de la Unión de ACh a los receptores, canales específicos se vuelven igualmente permeables a Na+ y K+. Este cambio de permeabilidad tiende a mover el potencial de membrana desde donde está inicialmente hacia un nuevo potencial de 0 mV.
¿Por qué el potencial de endplate normal nunca alcanza realmente 0 mV?, Una de las razones es que la secuencia de cambios de permeabilidad que subyacen al potencial de acción «ahogan» los cambios producidos por el PPE. Pero incluso si no se activara un potencial de acción, el PPE aún no alcanzaría 0 mV. Esto se debe a que los canales ACh son solo una pequeña fracción del número total de canales en las fibras musculares. Los canales K+ que dotan a una célula muscular de su potencial de reposo también están presentes. Su trabajo es tratar de mantener la célula en el potencial de reposo.,
el canal abierto por ACh es un miembro de una clase general de canales llamados canales controlados por ligando o receptores ionotrópicos. Como se ilustra en la figura 4.10, el sitio de enlace del transmisor es parte del propio canal. Como resultado de la Unión del transmisor al receptor (generalmente se necesitan dos moléculas), hay un cambio conformacional en la proteína que permite que una región de poro se abra y los iones fluyan hacia abajo sus gradientes electroquímicos. Los detalles adicionales del canal se presentan en el Capítulo 11.,
Pruebe sus conocimientos
- Pregunta 1
- A
- B
- C
- D
un potencial endplate en una célula del músculo esquelético podría en principio ser producido por una permeabilidad disminuida a cuál de los iones siguientes? (Supongamos que hay una permeabilidad inicial finita a cada uno de los iones enumerados a continuación y que los gradientes de concentración fisiológicos están presentes.):
A. Na+
B. Na+ y Ca2+
C. Ca2+
D., K+
un potencial endplate en una célula del músculo esquelético podría en principio ser producido por una permeabilidad disminuida a cuál de los iones siguientes? (Supongamos que hay una permeabilidad inicial finita a cada uno de los iones enumerados a continuación y que los gradientes de concentración fisiológicos están presentes.):
A. Na+ Esta respuesta es INCORRECTA.
un potencial de placa final es una despolarización que normalmente se produce por el aumento simultáneo de la permeabilidad a los iones de sodio y potasio., Si hubiera una disminución selectiva en la permeabilidad al sodio, tal disminución no conduciría a una despolarización. Más bien, en realidad conduciría a una hiperpolarización porque alfa en la ecuación de GHK se reduciría. La relación de permeabilidad a sodio y potasio se favorecería hacia la permeabilidad a potasio, desplazando el potencial de membrana hacia el potencial de equilibrio de potasio y produciendo una hiperpolarización.
B. Na+ y Ca2+
C. Ca2+
D., K+
un potencial endplate en una célula del músculo esquelético podría en principio ser producido por una permeabilidad disminuida a cuál de los iones siguientes? (Supongamos que hay una permeabilidad inicial finita a cada uno de los iones enumerados a continuación y que los gradientes de concentración fisiológicos están presentes.):
A. Na+
B. Na+ y Ca2+, Esta respuesta es INCORRECTA.si se disminuyera la permeabilidad al sodio y al calcio, sus consecuencias serían similares a las de la opción A. La disminución de la permeabilidad al sodio por sí sola tendería a hiperpolarizar la célula., Del mismo modo, la disminución de la permeabilidad al calcio también podría hiperpolarizar la célula. El potencial de equilibrio de calcio es un valor muy positivo, y si hubiera alguna permeabilidad tónica en reposo al calcio, esa permeabilidad contribuiría a una despolarización tónica del potencial de membrana. Por lo tanto, una disminución en la permeabilidad del calcio eliminaría ese efecto tónico despolarizante y daría lugar a una hiperpolarización.
C. Ca2+
D., K+
un potencial endplate en una célula del músculo esquelético podría en principio ser producido por una permeabilidad disminuida a cuál de los iones siguientes? (Supongamos que hay una permeabilidad inicial finita a cada uno de los iones enumerados a continuación y que los gradientes de concentración fisiológicos están presentes.):
A. Na+
B. Na+ y Ca2+
C. Ca2+, Esta respuesta es INCORRECTA.
una disminución de la permeabilidad al calcio por sí sola produciría, en todo caso, una hiperpolarización. Véase lógica de respuesta a la opción B.
D., K+
un potencial endplate en una célula del músculo esquelético podría en principio ser producido por una permeabilidad disminuida a cuál de los iones siguientes? (Supongamos que hay una permeabilidad inicial finita a cada uno de los iones enumerados a continuación y que los gradientes de concentración fisiológicos están presentes.):
A. Na+
B. Na+ y Ca2+
C. Ca2+
D. K+ Esta respuesta es CORRECTA!
una disminución en la permeabilidad de potasio conduciría a una despolarización similar a un potencial de placa terminal., Esto es así porque hay en reposo una permeabilidad tónica al potasio y al sodio. La alta permeabilidad al potasio tiende a mantener el potencial de membrana cerca del potencial de equilibrio de potasio. Si se disminuye esa permeabilidad en reposo, alfa en la ecuación de Goldman se convertiría en un valor mayor, moviendo el potencial de membrana un poco más cerca del potencial de equilibrio de sodio (es decir, una despolarización).