Introduction
La grande perspicacité de Darwin, et le principe unificateur de la biologie aujourd’hui, est que toutes les espèces sont liées les unes aux autres comme des Sœurs, des cousins et des parents éloignés dans un Les implications sont à couper le souffle; si nous pouvions voyager assez loin dans le temps, nous trouverions des ancêtres communs entre nous et tous les autres organismes vivants, des porcs-épics aux flamants roses en passant par les cactus., Notre famille évolutive immédiate est composée des hominoïdes, le groupe de primates qui comprend les « petits singes » (siamangs et gibbons) ainsi que les « grands singes » (chimpanzés, bonobos, gorilles et orangs-outans). Parmi les grands singes, nos plus proches parents sont les chimpanzés et les bonobos (Figure 1). Les archives fossiles, ainsi que les études de L’ADN humain et des singes, indiquent que les humains partageaient un ancêtre commun avec les chimpanzés et les bonobos il y a environ 6 millions d’années (mya)., Nous commençons cette discussion sur l’évolution de notre espèce en Afrique, vers la fin de la période géologique connue sous le nom de Miocène, juste avant que notre lignée ne diverge de celle des chimpanzés et des bonobos.
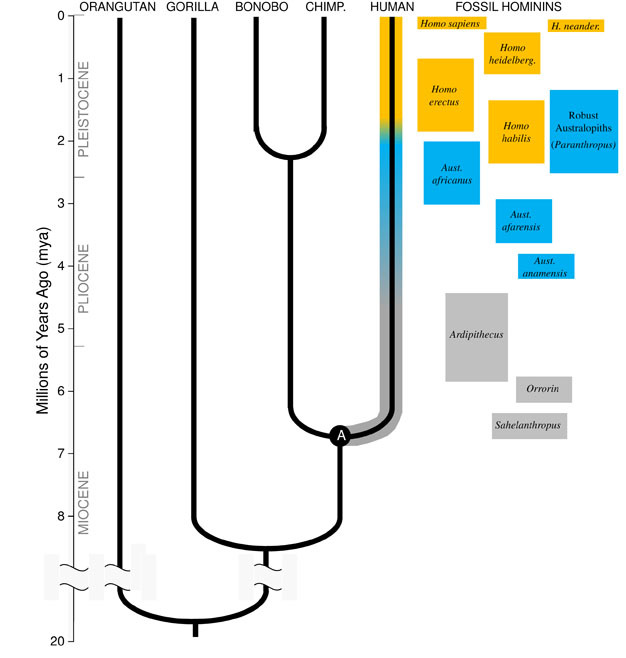
Miocène Origines de la Lignée des Hominiens
afin de comprendre l’évolution de toute espèce, nous devons d’abord établir ancestrales de l’etat: quelle sorte d’animal fait-il évoluer? Pour notre lignée, cela nécessite que nous essayions de reconstruire le dernier ancêtre commun des humains et des chimpanzés (marqué « A » sur la Figure 1). Le dernier ancêtre commun humain-chimpanzé (HC-LCA) est l’espèce à partir de laquelle la lignée des hominins et la lignée des chimpanzés & Bonobo ont divergé., Les hominines sont des espèces de notre branche de l’arbre hominoïde après la scission avec la lignée des chimpanzés & bonobo, y compris toutes les espèces éteintes et les branches latérales évolutives (Figure 1).
Il y avait une grande diversité d’espèces de singes au Miocène, avec des dizaines d’espèces connues à partir des archives fossiles en Afrique, en Europe et en Asie. Ces espèces ont varié dans leur anatomie et leur écologie, et il n’est pas clair qui, le cas échéant, des espèces fossiles découvertes jusqu’à présent représentent le HC-LCA (Kunimatsu et al. 2007; Young et MacLatchy, 2004)., Néanmoins, nous savons par des preuves fossiles et comparatives qu’il était beaucoup plus similaire aux singes vivants qu’aux humains vivants. Le HC-LCA aurait eu un cerveau et un corps de la taille d’un singe, avec des bras et des doigts relativement longs et un pied agrippant qui lui a permis de se nourrir dans les arbres. Les dents canines étaient probablement grandes et pointues, comme on l’a vu chez plusieurs hominoïdes du Miocène. De plus, les canines étaient probablement sexuellement dimorphes, les mâles ayant des canines beaucoup plus grandes que les femelles, comme on le voit chez les grands singes vivants et les fossiles du Miocène., Comme les singes vivants, il aurait marché en quadrupède (à quatre pattes) lorsqu’il était sur le sol, et son régime alimentaire aurait consisté presque entièrement en aliments végétaux, principalement des fruits et des feuilles.
Hominines précoces
des modifications de l’anatomie d’un singe sont perceptibles dans des fossiles d’hominoïdes du Miocène supérieur en Afrique. Certaines espèces d’hominoïdes de cette période présentent des traits typiques des humains mais ne sont pas visibles chez les autres singes vivants, ce qui conduit les paléoanthropologues à déduire que ces fossiles représentent les premiers membres de la lignée des hominines., Les premiers traits de type humain à apparaître dans les archives fossiles d’hominines sont la marche bipède et les canines plus petites et émoussées.
Les plus anciens hominins actuellement connus sont Sahelanthropus tchadensis du Tchad (Brunet et al. 2005) et Orrorin tugenensis du Kenya (Senut et coll. 2001). Sahelanthropus, daté entre 6 et 7 mya, est connu à partir d « un crâne en grande partie complet et d » autres restes fragmentaires. Sa taille de cerveau, 360cc, est dans la gamme observée chez les chimpanzés, et le crâne a une crête frontale massive, similaire en épaisseur aux gorilles mâles (Brunet et al. 2005)., Cependant, la position et l’orientation du foramen magnum, le trou dans la base du crâne à travers lequel passe la moelle épinière, suggère que Sahelanthropus se tenait et marchait bipède, avec sa colonne vertébrale maintenue verticalement comme chez les humains modernes plutôt qu’horizontalement comme chez les singes et autres quadrupèdes (Zollikofer et al. 2005). Orrorin est connu principalement à partir de fossiles post-crâniens, y compris un fémur partiel. La partie proximale du fémur présente des similitudes avec celles des humains modernes, suggérant que l’espèce était bipède (Pickford et al. 2002)., Aucun crâne D’Orrorin n’a été retrouvé, et sa morphologie crânienne et sa taille cérébrale sont donc incertaines. Chez Orrorin et Sahelanthropus, les dents canines des mâles sont plus grandes et plus pointues que chez les humains modernes, mais sont petites et émoussées par rapport aux canines des singes mâles. Cela suggère que le dimorphisme sexuel canin — et par extension, la compétition entre les mâles pour l’accès à l’accouplement des femelles-a été diminué chez ces premiers hominins par rapport aux grands singes.
de loin L’hominine primitive la plus connue est Ardipithecus ramidus, a 4.,Espèce vieille de 4 millions d’années D’Éthiopie, connue à partir d’un squelette presque complet ainsi que de nombreux autres restes dentaires et squelettiques (White et al. 2009). Ar. ramidus et une plus ancienne, espèces apparentées connues à partir de restes fragmentaires, Ar. kadabba (5.8-5.2 mya), ont des canines réduites similaires à celles d’Orrorin et de Sahelanthropus. Le crâne de l’Ar. ramidus est plutôt semblable à un singe et largement similaire à celui de Sahelanthropus, avec un petit cerveau de la taille d’un chimpanzé de 300-350cc (Figure 2). Le squelette Postcrânien D’Ardipithecus est intrigant., Bien que mal fragmenté, le bassin récupéré révèle une morphologie très différente de celle des singes vivants, avec une forme plus courte et plus en forme de bol qui suggère fortement Qu’Ardipithecus marchait bipède; ceci est cohérent avec la position du foramen magnum, qui suggère une posture verticale. Cependant, ses longs membres antérieurs et ses doigts et son premier orteil divergent et saisissant (hallux) suggèrent Qu’Ardipithecus a passé une grande partie de son temps dans les arbres. L’impression générale est celle d’une espèce en grande partie arboricole qui marchait bipède chaque fois qu’elle s’aventurait au sol.,
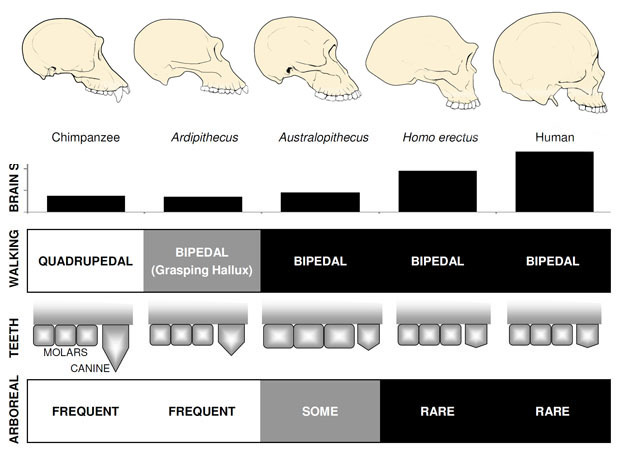
Australopithecus
Autour de 4mya nous trouvons les premiers membres du genre Australopithecus, les homininés qui ont été adepte terrestre bipèdes, mais a continué à utiliser les arbres pour la nourriture et la protection. Les premiers spécimens D’Australopithèques ont été découverts en Afrique du Sud en 1924 (Dart, 1925), et les efforts de recherche au cours des huit décennies suivantes ont produit des centaines de fossiles de plusieurs espèces sur des sites partout en Afrique orientale et australe., Nous savons maintenant que Australopithecus était un genre très réussi qui a persisté pendant près de trois millions d’années (Figure 1).
Les espèces D’Australopithèques les plus connues sont A. afarensis (3.6-2.9 mya) d’Afrique de l’Est et A. africanus (3.2–2.0 mya) d’Afrique du Sud., Le bassin et le membre inférieur de ces espèces indiquent clairement qu’ils étaient complètement bipèdes: le bassin est court et en forme de bol, amenant les muscles fessiers sur le côté du corps, comme chez l’homme moderne, pour la stabilisation du tronc pendant la bipédie, et le premier orteil est aligné avec les autres orteils (Ward, 2002; Harcourt-Smith et Aiello, 2004). Le pied de L’Australopithèque pourrait même avoir eu une arche de type humain, d’après l’analyse des métatarses et des empreintes fossilisées de Laetoli (Ward et al. 2011)., Néanmoins, par rapport aux humains modernes, les avant-bras étaient longs et les doigts et les orteils étaient longs et quelque peu courbés, ce qui suggère que les australopithèques utilisaient régulièrement les arbres pour se nourrir et peut-être comme refuge contre les prédateurs la nuit. Cette stratégie arboricole mixte terrestre & aurait bien servi ces espèces dans les milieux mixtes boisés et savanes qu’elles habitaient.
La Taille du cerveau chez les australopithèques variait entre 390 et 515 CC, semblable à celle des chimpanzés et des gorilles (Falk et al., 2000), suggérant que les capacités cognitives étaient largement similaires à celles des singes vivants (Figure 2). Chez les australopithèques, la taille du corps était plutôt petite et sexuellement dimorphique, environ 30 kg pour les femelles et 40 kg pour les mâles (McHenry, 1992). Ce niveau de dimorphisme ne se reflète pas dans les canines, qui étaient petites, émoussées et monomorphes comme chez les hominines antérieures.
Contrairement aux canines, les dents molaires des australopithèques étaient beaucoup plus grandes que celles des hominins antérieurs et avaient un émail plus épais. Cela suggère que leur régime alimentaire comprenait des aliments végétaux durs et de mauvaise qualité qui nécessitaient une mastication puissante pour être traités., Un sous-groupe D’Australopithèques, connu sous le nom d’australopithèques « robustes » (souvent étiquetés par un genre distinct Paranthropus) en raison de leurs énormes dents et de leurs muscles à mâcher, a poussé cette adaptation à l’extrême. La plupart des espèces D’Australopithèques ont été éteintes par 2 mya, mais certaines formes robustes ont persisté jusqu’à environ 1.2 mya en Afrique de l’Est et du Sud.
le genre Homo
Les premiers fossiles de notre propre genre, Homo, se trouvent en Afrique de l’Est et datés de 2,3 mya (Kimbel et al. 1997)., Ces premiers spécimens sont similaires en taille du cerveau et du corps à Australopithecus, mais montrent des différences dans leurs dents molaires, suggérant un changement de régime alimentaire. En effet, à au moins 1,8 mya, les premiers membres de notre genre utilisaient des outils de Pierre primitifs pour boucher les carcasses d’animaux, ajoutant de la viande et de la moelle osseuse riches en énergie à leur régime alimentaire à base de plantes.
Le membre le plus ancien du genre Homo, H. habilis (2.3–1.4 mya) se trouve en Afrique de l’Est et est associé à des os d’animaux massacrés et à de simples outils en pierre (Blumenschine et al. 2003). Son descendant le plus redoutable et le plus répandu, H., erectus, se trouve dans toute L’Afrique et L’Eurasie et a persisté de 1,9 mya à 100 kya, et peut-être même plus tard (Anton, 2003). Comme les humains modernes, H. erectus n’avait pas les adaptations des membres antérieurs pour l’escalade observées chez L’Australopithèque (Figure 2). Son expansion mondiale suggère que H. erectus était écologiquement flexible, avec la capacité cognitive de s’adapter et de prospérer dans des environnements très différents. Sans surprise, c’est avec H. erectus que nous commençons à voir une augmentation majeure de la taille du cerveau, jusqu’à 1 250 cc pour les spécimens asiatiques ultérieurs (Anton, 2003). La taille molaire est réduite en H., erectus par rapport à Australopithecus, reflétant son régime alimentaire plus doux et plus riche.
vers 700 kya, et peut-être plus tôt, H. erectus en Afrique a donné naissance à H. heidelbergensis, une espèce qui nous ressemble beaucoup en termes de proportions corporelles, d’adaptations dentaires et de capacités cognitives (Rightmire, 2009). H. heidelbergensis, souvent appelé Homo sapiens « archaïque », était un chasseur de gros gibier actif, produisait des outils sophistiqués de style Levallois et, Vers au moins 400 kya, avait appris à maîtriser le feu (Roebroeks et Villa, 2011). Les Néandertaliens (H., neanderthalensis), des hominins adaptés au froid avec un physique robuste, des comportements complexes et un cerveau de taille similaire au nôtre, auraient évolué à partir des populations de H. heidelbergensis en Europe d’au moins 250 kya (Rightmire, 2008; Hublin, 2009).
des preuves fossiles et ADN suggèrent que notre propre espèce, H. sapiens, a évolué en Afrique 200 kya (Relethford, 2008; Rightmire, 2009), probablement à partir de H. heidelbergensis. La sophistication comportementale accrue de H., sapiens, comme l’indiquent nos grands cerveaux (1400 cc) et les preuves archéologiques d’un ensemble d’outils plus large et de techniques de chasse intelligentes, ont permis à notre espèce de s’épanouir et de se développer sur le continent africain. À 100kya, notre espèce s’est répandue en Eurasie, pour finalement s’étendre à travers le monde entier en Australie et dans les Amériques (DiGiorgio et al. 2009). En cours de route, nos espèces ont déplacé d’autres hominins qu’elles ont rencontrés, y compris les Néandertaliens en Europe et des formes similaires en Asie. (Notez que tous ne sont pas d’accord avec cette interprétation des données, voir Tryon et Bailey)., Les études de L’ADN ancien extrait de fossiles de Néandertal suggèrent que notre espèce pourrait avoir parfois intercalé avec eux (Green et al., 2010). Notre impact mondial croissant se poursuit aujourd’hui, alors que les innovations culturelles telles que l’agriculture et l’urbanisation façonnent le paysage et les espèces qui nous entourent.
résumé
l’évolution de notre espèce à partir d’un ancêtre semblable à un singe du Miocène a été un processus complexe. Notre lignée est pleine de branches latérales et d’impasses évolutives, avec des espèces comme les australopithes robustes qui ont persisté pendant plus d’un million d’années avant de disparaître., Certains traits humains, comme la bipédie, ont évolué très tôt, tandis que d’autres, comme les gros cerveaux, n’ont évolué que relativement récemment. D’autres traits encore, comme la taille molaire, ont évolué dans une direction pour être repoussés plus tard par des pressions écologiques changeantes. Plutôt qu’un navire puissant traçant une route droite vers une destination prédéterminée, l’évolution de notre lignée-en fait, de la lignée de n’importe quelle espèce — correspond à l’image d’un canot de sauvetage ballotté par les mers changeantes du changement environnemental, de la chance génétique et du hasard géologique. On se demande où les six prochains millions d’années pourraient nous mener.