introduktion
Darwins stora insikt, och den förenande principen om biologi idag är att alla arter är relaterade till varandra som systrar, kusiner och avlägsna släktingar i ett stort släktträd av livet. Konsekvenserna är hisnande; om vi kunde resa tillbaka tillräckligt långt i tiden, skulle vi hitta gemensamma förfäder mellan oss själva och alla andra levande organismer, från piggsvin till Flamingos till Kaktusar., Vår närmaste evolutionära familj består av hominoiderna, den grupp primater som innehåller de ” mindre aporna ”(siamangs och gibbons) samt de” stora aporna ” (schimpanser, bonobos, gorillor och orangutanger). Bland de stora aporna är våra närmaste släktingar schimpanserna och bonobos (Figur 1). Den fossila skivan, tillsammans med studier av humant och ape-DNA, indikerar att människor delade en gemensam förfader med schimpanser och bonobos någon gång omkring 6 miljoner år sedan (mya)., Vi börjar denna diskussion om vår arts utveckling i Afrika, nära slutet av den geologiska tidsperiod som kallas miocen, strax innan vår härstamning avvikit från den av schimpanser och bonobos.
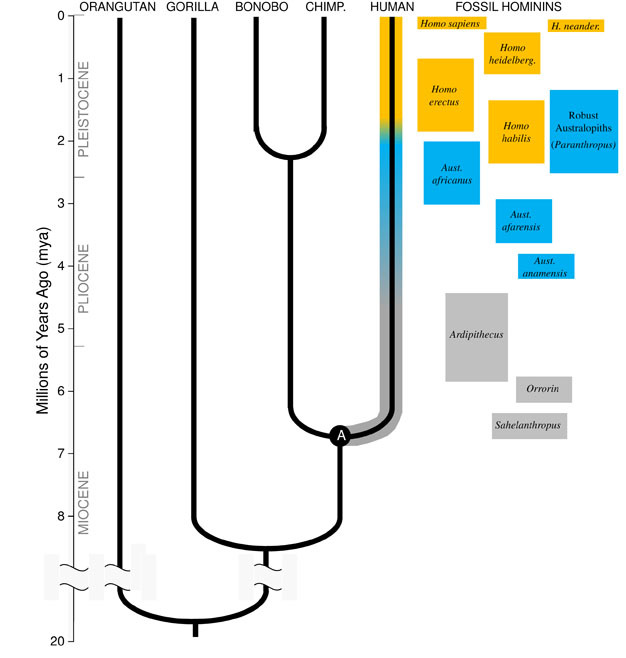
miocen Ursprung Hominin härstamning
för att förstå utvecklingen av någon art, måste vi först fastställa dess förfäders tillstånd: vilken typ av djur utvecklades det från? För vår härstamning kräver detta att vi försöker rekonstruera den sista gemensamma förfadern till människor och schimpanser (märkt ”A” i Figur 1). Den Human-schimpans sista gemensamma förfader (HC-LCA) är den art från vilken hominin härstamning och schimpans & bonobo härstamning divergerade., Homininer är arter på vår gren av hominoidträdet efter splittringen med schimpansen & bonobo-linjen, inklusive alla utdöda arter och evolutionära sidogrenar (Figur 1).
det fanns en stor mångfald av aparter i miocen, med dussintals arter kända från fossila rekord över Afrika, Europa och Asien. Dessa arter varierade i deras anatomi och ekologi, och det är inte klart vilken, om någon, av de fossila arter som hittills upptäckts representerar HC-LCA (Kunimatsu et al. 2007, Unga och MacLatchy, 2004)., Ändå vet vi från fossila och jämförande bevis att det var mycket mer lik levande apor än levande människor. HC-LCA skulle ha haft en apa-storlek hjärna och kropp, med relativt långa armar och fingrar och en gripande fot som gjorde det möjligt att foder i träden. Hundtänderna var förmodligen stora och skarpa, vilket ses i flera Miocena hominoider. Dessutom var kaninerna förmodligen sexuellt dimorfa, med män som hade mycket större hundar än kvinnor, vilket ses bland de levande stora aporna och Miocenfossilen., Liksom levande apor skulle det ha gått kvadrupedally (på alla fyra) när på marken, och dess kost skulle ha bestod nästan helt av vegetabiliska livsmedel, främst frukt och blad.
tidiga homininer
förändringar från en apliknande anatomi kan urskiljas i hominoidfossil från den sena miocen i Afrika. Vissa hominoid-arter från denna period uppvisar egenskaper som är typiska för människor men ses inte i de andra levande aporna, vilket leder paleoantropologer att dra slutsatsen att dessa fossil representerar tidiga medlemmar av hominin-linjen., De första mänskliga-liknande egenskaper som visas i homin fossil posten är tvåbent promenader och mindre, trubbiga hundar.
Den äldsta hominins känd för närvarande är Sahelanthropus tchadensis från Tchad (Brunet m.fl. 2005) och Orrorin tugenensis från Kenya (Senut et al. 2001). Sahelanthropus, daterat till mellan 6 och 7 mya, är känt från en till stor del komplett skalle och några andra fragmentariska rester. Dess hjärnstorlek, 360cc, ligger inom intervallet sett i schimpanser, och skallen har en massiv panna ås, liknande i tjocklek till manliga gorillor (Brunet et al. 2005)., Emellertid, positionen och orienteringen av foramen magnum, hålet i basen av skallen genom vilken ryggmärgen passerar, tyder på att Sahelanthropus stod och gick bipedally, med dess ryggrad hålls vertikalt som i moderna människor snarare än horisontellt som i apor och andra kvadrupeder (Zollikofer et al. 2005). Orrorin är främst känd från postkraniella fossil, inklusive en partiell lårben. Den proximala delen av lårbenet visar likheter med moderna människor, vilket tyder på att arten var bipedal (Pickford et al. 2002)., Inga skallar av Orrorin har återhämtats, och så är dess kranialmorfologi och hjärnstorlek osäker. I både Orrorin och Sahelantropus är hundtänderna hos män större och mer spetsiga än hos moderna människor, men är små och trubbiga jämfört med hundar av manliga apor. Detta tyder på att hund sexuell dimorfism — och i förlängningen konkurrens bland män för parning tillgång till kvinnor-minskade i dessa tidiga homininer jämfört med de stora aporna.
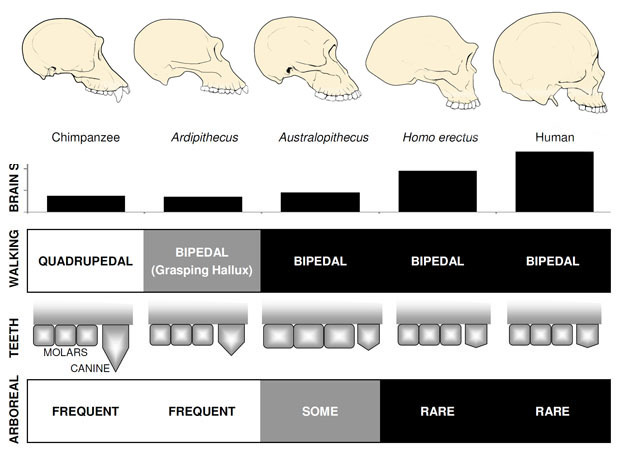
den överlägset mest kända tidiga hominin är Ardipithecus ramidus, en 4.,4 miljoner år gamla arter från Etiopien, som är känd från en nästan komplett skelett samt många andra tand-och skelettrester (White et al. 2009). Ar. ramidus och en äldre, relaterade arter kända från fragmentariska rester, Ar. kadabba (5.8–5.2 mya), har minskat hundar som liknar Orrorin och Sahelanthropus. Skallen av Ar. ramidus är ganska apliknande och liknar i stort sett Sahelanthropus, med en liten schimpansstorlek av 300-350cc (Figur 2). Ardipithecus postcranial skelett är spännande., Även om dåligt fragmenterad, bäckenet återhämtat sig avslöjar en morfologi helt annorlunda än den hos levande apor, med en kortare, mer skålliknande form som starkt föreslår Ardipithecus gick bipedally; detta överensstämmer med foramen magnum position, vilket tyder på en upprätt hållning. Men dess långa framben och fingrar och dess divergerande, gripande första tå (hallux) föreslår Ardipithecus tillbringade mycket av sin tid i träden. Helhetsintrycket är av en till stor del arboreal art som gick tvåbent när den vågade till marken.,
Australopithecus
runt 4mya hittar vi de tidigaste medlemmarna av släktet Australopithecus, homininer som var adept terrestrial bipeds men fortsatte att använda träden för mat och skydd. De första exemplaren av Australopithecus upptäcktes i Sydafrika 1924 (Dart, 1925), och forskningsinsatser under de efterföljande åtta decennierna har producerat hundratals fossil från flera arter på platser över hela östra och Södra Afrika., Vi vet nu att Australopithecus var ett mycket framgångsrikt släkte som kvarstod i nästan tre miljoner år (Figur 1).
de mest kända Australopithecus arterna är A. afarensis (3.6–2.9 mya) från Östafrika och A. africanus (3.2–2.0 mya) från Sydafrika., Bäckenet och underbenet av dessa arter indikerar tydligt att de var helt bipedal: bäckenet är kort och skålformad, vilket ger de gluteala musklerna runt sidan av kroppen, som i moderna människor, för stammen stabilisering under bipedalism, och den första tånen är i linje med de andra tårna (Ward, 2002; Harcourt-Smith och Aiello, 2004). Australopithecus foten kan till och med ha haft en mänsklig-liknande båge, baserad på analys av metatarsaler och fossiliserade Laetoli fotspår (Ward et al. 2011)., Men jämfört med moderna människor var underarmarna långa och fingrarna och tårna var långa och något böjda, vilket tyder på att Australopithecus regelbundet använde träden till foder och kanske som en tillflykt från rovdjur på natten. Denna blandade markbundna& arboreala strategi skulle ha tjänat dessa arter väl i den blandade skogsmark och savannah miljöer de bebodda.
hjärnstorlek i Australopithecus varierade mellan 390 och 515cc, liknande schimpanser och gorillor (Falk et al., 2000), vilket tyder på kognitiva förmågor, liknade i stort sett levande apor (Figur 2). Kroppsstorlek i Australopithecus var ganska liten och sexuellt dimorfisk, ca 30kg för kvinnor och 40kg för män (McHenry, 1992). Denna nivå av dimorfism återspeglas inte i hundar, som var små, trubbiga och monomorfa som i tidigare homininer.
Till skillnad från hundar, molar tänder i Australopithecus var mycket större än tidigare homininer, och hade tjockare emalj. Detta tyder på att deras kost innehöll hårda, lågkvalitativa växtfoder som krävde kraftfull tuggning för att bearbeta., En undergrupp av Australopithecus, känd som ”robusta” australopiths (ofta märkt av ett separat släkte Paranthropus) på grund av sina enorma tänder och tuggmuskler, tog denna anpassning till det yttersta. De flesta Australopithecus arter utrotades av 2 mya, men vissa robusta former kvarstod tills ca 1,2 mya i östra och Sydafrika.
släktet Homo
de tidigaste fossilerna i vårt eget släkte Homo finns i Östafrika och dateras till 2,3 mya (Kimbel et al. 1997)., Dessa tidiga exemplar är liknande i hjärnan och kroppsstorlek till Australopithecus, men visar skillnader i sina molar tänder, vilket tyder på en förändring i kost. Faktum är att åtminstone 1,8 mya använde tidiga medlemmar av vårt släkt primitiva stenverktyg för att slakta djurkroppar och lägga till energirik kött och benmärg till sin växtbaserade diet.
den äldsta medlemmen av släktet Homo, H. habilis (2.3–1.4 mya) finns i Östafrika och är associerad med slaktade djurben och enkla stenverktyg (Blumenschine et al. 2003). Dess mer formidabla och utbredda ättling, H., erectus, finns i hela Afrika och Eurasien och kvarstod från 1,9 mya till 100 kya, och kanske till och med senare (Anton, 2003). Liksom moderna människor saknade H. erectus forelimb-anpassningarna för klättring som ses i Australopithecus (Figur 2). Dess globala expansion tyder på att H. erectus var ekologiskt flexibel, med kognitiv förmåga att anpassa sig och trivas i väldigt olika miljöer. Inte överraskande är det med H. erectus att vi börjar se en stor ökning av hjärnstorleken, upp till 1,250 cc för senare asiatiska exemplar (Anton, 2003). Molar storlek reduceras i H., erectus i förhållande till Australopithecus, vilket återspeglar dess mjukare, rikare diet.
omkring 700 kya, och kanske tidigare, H. erectus i Afrika gav upphov till H. heidelbergensis, en art som oss väldigt mycket när det gäller kroppsproportioner, tandanpassningar och kognitiv förmåga (Rightmire, 2009). H. heidelbergensis, som ofta refereras till som en ”ålderdomlig” Homo sapiens, var en aktiv big-game hunter, som produceras sofistikerade Levallois stil verktyg, och med minst 400 kya hade lärt sig att kontrollera eld (Roebroeks och Villa, 2011). Neandertalarna (H., neanderthalensis), kall-anpassad hominins med kraftiga physiques, komplexa beteenden, och hjärnor liknande i vår storlek, tros ha utvecklats från H. heidelbergensis populationer i Europa med minst 250 kya (Rightmire, 2008; Hublin, 2009).
fossila och DNA bevis tyder på vår egen art, H. sapiens, utvecklats i Afrika 200 kya (Relathford, 2008; Rightmire, 2009), förmodligen från H. heidelbergensis. Den ökade beteendemässiga sofistikeringen av H., sapiens, som indikeras av våra stora hjärnor (1,400 cc) och arkeologiska bevis på ett bredare verktyg och smarta jakttekniker, gjorde det möjligt för våra arter att blomstra och växa på den afrikanska kontinenten. Av 100kya spillde vår art i Eurasien, så småningom expanderar över hela världen till Australien och Amerika (DiGiorgio et al. 2009). Längs vägen våra arter förskjutit andra homininer de stött på, inklusive Neanderthals i Europa och liknande former i Asien. (Observera att inte alla håller med denna tolkning av data, se Tryon och Bailey)., Studier av gammalt DNA extraherat från Neanderthal fossil tyder på att våra arter kan ha ibland interbred med dem (Green et al., 2010). Vår ökande globala påverkan fortsätter idag, eftersom Kulturella innovationer som jordbruk och urbanisering formar landskapet och arten omkring oss.
sammanfattning
utvecklingen av vår art från en apliknande miocen förfader var en komplex process. Vår härstamning är full av sidogrenar och evolutionära återvändsgränder, med arter som de robusta australopiterna som kvarstod i över en miljon år innan de bleknar bort., Vissa mänskliga egenskaper, som bipedalism, utvecklades väldigt tidigt, medan andra, som stora hjärnor, inte utvecklades förrän relativt nyligen. Fortfarande andra egenskaper, som molarstorlek, utvecklades i en riktning bara för att skjutas tillbaka senare genom att ändra ekologiska tryck. I stället för ett kraftfullt fartyg som kartlägger en rak kurs mot något förutbestämt resmål passar utvecklingen av vår härstamning-faktiskt av någon Arts släkt — bilden av en livbåt som kastas om av de skiftande Haven av miljöförändring, genetisk lycka och geologisk chans. Man undrar vart de kommande sex miljoner åren kan ta oss.