Figura 4.1 (a se vedea mărită)
prin Urmare, o înțelegere de acest synapse duce la o înțelegere a altora. Prin urmare, vom discuta mai întâi procesul de transmitere sinaptică la joncțiunea neuromusculară scheletică.,
caracteristicile joncțiunii sinaptice la joncțiunea neuromusculară sunt prezentate în figura din stânga. Fibrele musculare scheletice sunt inervate de neuronii motori ale căror corpuri celulare sunt situate în Cornul ventral al măduvei spinării. Regiunea terminală a axonului dă naștere unor procese foarte fine care se desfășoară de-a lungul celulelor musculare scheletice. De-a lungul acestor procese sunt structuri specializate cunoscute sub numele de sinapse. Sinapsa special făcut între un neuron motor spinal și celule musculare scheletice este numit endplate motor din cauza structurii sale specifice.,sinapsa la joncțiunea neuromusculară are trei caracteristici caracteristice ale sinapselor chimice din sistemul nervos. În primul rând, există o separare distinctă între membrana presinaptică și postsynaptică. Spațiul dintre cele două este cunoscut sub numele de cleft sinaptic. Spațiul ne spune că trebuie să existe un mecanism intermediar de semnalizare între neuronul presinaptic și neuronul postsinaptic pentru a avea fluxul de informații prin fanta sinaptică. În al doilea rând, există o densitate mare caracteristică a veziculelor sferice mici., Aceste vezicule sinaptice conțin substanțe neurotransmițătoare. Sinapsele sunt, de asemenea, asociate cu o densitate mare de mitocondrii. În al treilea rând, în majoritatea cazurilor, există o îngroșare caracteristică a membranei postsinaptice, care se datorează cel puțin parțial faptului că membrana postsinaptică are o densitate mare de receptori specializați care leagă substanțele emițătoare chimice eliberate din neuronul presinaptic. Detalii suplimentare privind caracteristicile morfologice ale joncțiunilor sinaptice sunt furnizate în capitolele 8 și Capitolul 10.
4.,2 Fiziologie a Transmisiei Sinaptice la nivelul plăcii Neuromusculare
Figura 4.2
figura din dreapta ilustrează într-un mod foarte schematic modul cum este posibil de a studia fiziologia transmisiei sinaptice la osos joncțiunii neuromusculare în detaliu. O bucată de mușchi și nervul atașat sunt plasate într-o mică cameră experimentală umplută cu o soluție de sonerie adecvată. Potențialul de repaus al celulei musculare este înregistrat cu un microelectrod. Electrozii sunt de asemenea plasați pe suprafața axonului nervos., Șocurile electrice scurte determină inițierea potențialelor de acțiune, care se propagă la terminalul sinaptic. figura de mai jos ilustrează două tipuri de modificări potențiale care au fost înregistrate într-un astfel de preparat nervos-muscular izolat. Experimentul ilustrează, de asemenea, proprietățile unui medicament puternic, curare, care sa dovedit a fi foarte util în studierea procesului de transmitere sinaptică la joncțiunea neuromusculară scheletică. Partea A ilustrează secvența modificărilor potențiale înregistrate în celula musculară ca urmare a stimulării axonului motor., Săgeata indică momentul în care șocul este livrat axonului motorului. Rețineți că există o perioadă de repaus după șoc. Întârzierea se datorează timpului necesar pentru ca potențialul de acțiune din axonul motorului să se propage de la locul său de inițiere. După întârziere, există două tipuri de potențiale înregistrate în celula musculară. În primul rând, există un potențial relativ lent în schimbare, care va fi punctul central al următoarei discuții., Dacă acest potențial inițial lent este suficient de mare, așa cum este în mod normal în celulele musculare scheletice, un al doilea potențial, un potențial de acțiune, este generat în celula musculară.
figura 4.3
potențialele de acțiune în celulele musculare scheletice se datorează mecanismelor ionice similare cu cele discutate anterior. Mai exact, există o schimbare dependentă de tensiune a permeabilității Na+ urmată de o creștere întârziată a permeabilității K+. (Cu toate acestea, pentru celulele musculare netede și celulele musculare cardiace, mecanismele ionice sunt diferite.,)
evenimentul de bază care declanșează potențialul de acțiune poate fi dezvăluit profitând de curare, o otravă săgeată folosită de unii indieni sud-americani. O doză mică de curare (Partea B) reduce evenimentul de bază, dar nu este încă suficient de redusă pentru a scădea sub prag. Dacă se administrează o doză ceva mai mare de curare (Partea C), evenimentul de bază lent devine subthreshold. Semnalul de bază este cunoscut sub numele de potențialul plăcii de capăt (PPE), deoarece este o schimbare potențială înregistrată la placa de capăt a motorului. În general, este cunoscut ca un potențial postsynaptic excitator (EPSP)., Curare blochează potențialul plăcii de capăt, deoarece este un inhibitor competitiv al acetilcolinei (ACh), transmițătorul eliberat la terminalul presinaptic. Curare nu blochează conductanța na + dependentă de tensiune sau conductanța k+ dependentă de tensiune care stă la baza potențialului de acțiune al mușchilor. Curare afectează stimulul (EPSP) care duce în mod normal la inițierea potențialului de acțiune musculară. Un animal care este otrăvit cu curare se va asfixia deoarece procesul de transmitere neuromusculară la mușchii respiratori este blocat., în mod normal, magnitudinea potențialului plăcii finale este destul de mare. Într-adevăr, amplitudinea potențialului plăcii finale este de aproximativ 50 mV, dar este nevoie doar de aproximativ 30 mV pentru a atinge pragul. Extra 20 mV se numește factorul de siguranță. Prin urmare, chiar dacă potențialul plăcii finale ar deveni ceva mai mic (de exemplu, 40 MV în amplitudine) din cauza oboselii, PPE ar atinge pragul și relația unu-la-unu dintre un potențial de acțiune în axonul motor și un potențial de acțiune în celula musculară ar fi păstrat.
figura 4.4
4.,3 propagarea PPE
care sunt proprietățile PPE și cum se compară cu proprietățile potențialului de acțiune?
este potențialul plăcii de capăt datorită unei modificări dependente de tensiune în permeabilitățile Na+ și K+ precum potențialul de acțiune?
este potențialul plăcii de capăt propagat într-un mod complet sau nimic ca potențialul de acțiune? figura din stânga ilustrează un experiment care examinează propagarea potențialului plăcii de capăt. Fibra musculară este trasă în țeapă în mod repetat cu electrozi la intervale de 1 mm., (Rețineți că potențialul plăcii de capăt este mic, deoarece acest experiment se face în prezența unei concentrații scăzute de curare, astfel încât potențialul plăcii de capăt poate fi înregistrat fără complicațiile declanșării unui potențial de acțiune.) Potențialul plăcii de capăt nu este propagat într-un mod total sau nimic. Se răspândește de-a lungul mușchiului, dar face acest lucru cu decrementare., Astfel, spread-ul de la placa motorie potențial de la site-ul său de deschidere către alte site-uri de-a lungul celulei musculare apare în mod pasiv și cu decrementare, doar ca subthreshold potențialul de a schimba într-o porțiune de axon se răspândește de-a lungul axonului, sau doar ca o schimbare în temperatură la un moment dat pe o tija de metal se răspândește de-a lungul tijei.
4.4 Prezentare generală a Secvenței de Evenimente de Fond al sistemului PPE
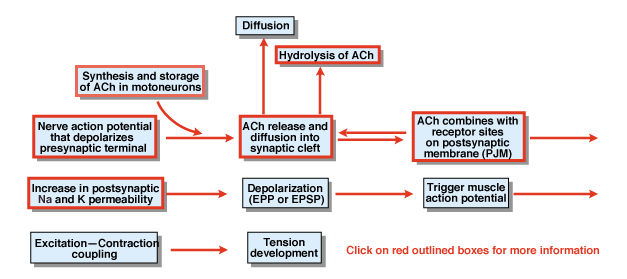
Figura 4.5
care sunt alte etape în procesul de chimice transmisiei sinaptice? Figura 4.,5 oferă o imagine de ansamblu. Un potențial de acțiune nervoasă care este inițiat în corpul celular al unui neuron motor spinal propagă rădăcinile ventrale și, în cele din urmă, invadează terminalele sinaptice ale neuronilor motori. Ca urmare a potențialului de acțiune, transmițătorul chimic acetilcolină (ACh) este eliberat în fanta sinaptică. ACh difuzează peste cleftul sinaptic și se leagă de receptorii specifici de pe membrana postsynaptică sau postjuncțională. Legarea ACh la receptorii săi produce o schimbare conformațională într-un canal membranar care este permeabil în mod specific atât la Na+ cât și la K+., Ca urmare a creșterii permeabilității Na+ și K+, există o depolarizare a membranei postsynaptice. Această depolarizare se numește potențialul plăcii finale sau, în general, EPSP. Dacă EPSP este suficient de mare, așa cum este în mod normal la joncțiunea neuromusculară, aceasta duce la inițierea unui potențial de acțiune în celula musculară. Potențialul de acțiune inițiază procesul de cuplare a contracției excitației și dezvoltarea tensiunii. Durata potențialului plăcii finale este de aproximativ 10 msec. doi factori controlează durata EPSP la joncțiunea neuromusculară., În primul rând, ACh este îndepărtat prin difuzie. În al doilea rând, o substanță din cleftul sinaptic, numită acetilcolinesterază (AChE), hidrolizează sau descompune ACh. AChE este una dintre cele mai eficiente enzime cunoscute. O singură moleculă de AChE poate hidroliza 600.000 de molecule de ACh pe minut.
4.5 Rolul de Durere
Figura 4.6
O familie importantă de substanțe, dintre care unul este neostigmina, inhibă acțiunea de Durere. Neostigmina blochează acțiunea AChE și, prin urmare, face potențialul plăcii finale mai mare și mai lung în durată., Această cifră ilustrează două potențiale endplate. Unul a fost înregistrat în soluție salină și curare și un al doilea înregistrat după ce neostigmina a fost adăugată la soluție. (Se adaugă Curare astfel încât proprietățile PPE să poată fi studiate fără a declanșa un potențial de acțiune în celula musculară.) După aplicarea neostigminei, potențialul plăcii de capăt este mult mai mare și mai lung. 4.6 a miastenia Gravis miastenia gravis este asociată cu slăbiciune musculară severă din cauza scăderii numărului de receptori de acetilcolină din celula musculară., Dacă potențialul plăcii de capăt este mai mic, potențialul plăcii de capăt nu va atinge pragul. Dacă nu atinge pragul, nu va exista potențial de acțiune în celula musculară și nici o contracție a mușchiului, ceea ce provoacă slăbiciune musculară. Neostigmina și alți inhibitori ai AChE sunt utilizați pentru a trata pacienții cu miastenia gravis. Acești agenți fac ca cantitatea de acetilcolină care este eliberată să ajungă mai eficient la receptorii acetilcolinei rămași.
4.,6b agenți nervoși
deși inhibitorii AChE au o valoare terapeutică importantă, unii inhibitori au fost și sunt încă folosiți ca otrăvuri. Unii inhibitori de durere, cum ar fi Soman și Sarin formează un bloc destul de ireversibil de durere. Acest bloc duce la niveluri extreme de ACh în fanta sinaptică. Persoanele atât de otrăvite mor din convulsii și spasticitate musculară, inclusiv mușchii respiratori.
figura 4.7
4.,7 iontoforeza ACh
iontoforeza este o tehnică interesantă care poate fi utilizată pentru a testa în continuare ipoteza că ACh este substanța neurotransmițătoare la joncțiunea neuromusculară. Dacă ACh este emițătorul care este eliberat de această sinapsă, s-ar prezice că ar trebui să fie posibilă înlocuirea aplicării artificiale a emițătorului pentru eliberarea normală a emițătorului. Deoarece ACh este o moleculă încărcată pozitiv, ea poate fi forțată să iasă dintr-un microelectrod pentru a simula eliberarea ACh de la un terminal presinaptic.
Figura 4.,8
într-adevăr, cantități minute de ACh pot fi aplicate în vecinătatea joncțiunii neuromusculare. Figura 4.8 compară un PPE produs prin stimularea axonului motor și răspunsul la ejecțiile ACh. Schimbarea potențială pare aproape identică cu potențialul plăcii finale produs de eliberarea normală a ACh. Acest experiment oferă suport experimental pentru conceptul că ACh este transmițătorul natural la această sinapsă.,răspunsul la ejecția ACh are și alte proprietăți interesante care sunt în concordanță cu natura colinergică a sinapsei la joncțiunea neuromusculară scheletică. Neostigmina face ca răspunsul la iontoforeza ACh să fie mai lung și mai mare. Curare reduce răspunsul deoarece concurează cu legarea normală a ACh. Dacă ACh este ejectat în celula musculară, nu se întâmplă nimic deoarece receptorii pentru acetilcolină nu sunt în interior; ele sunt în exteriorul celulei musculare., Aplicarea acetilcolinei în regiuni ale mușchiului departe de placa de capăt nu produce nici un răspuns, deoarece receptorii pentru ACh sunt concentrați în regiunea sinaptică. pentru a vă testa înțelegerea până acum, luați în considerare modul în care un agent precum TTX ar afecta generarea atât a unui PPE, cât și a răspunsului unei fibre musculare la aplicarea iontoforetică a ACh? TTX nu are niciun efect asupra răspunsului la ACh, dar blochează PPE. Motivul pentru care răspunsul la ACh nu este afectat este clar, dar mulți se așteaptă ca, dacă nu există niciun efect aici, să nu existe nici un efect asupra PPE., Tetrodotoxina nu afectează legarea acetilcolinei de receptori și, prin urmare, nu va afecta răspunsul la aplicarea directă a ACh. Cu toate acestea, tetrodotoxina va afecta capacitatea unui potențial de acțiune de a fi provocat în axonul motor. Dacă nu se poate obține un potențial de acțiune în axonul motorului, acesta nu poate provoca eliberarea emițătorului. Astfel, tetrodotoxina ar elimina total PPE. Blocul nu s-ar datora unui bloc de receptori ACh, ci mai degrabă unui bloc cu un pas înainte de eliberarea emițătorului.
4.,8 mecanisme ionice ale PPE
Bernard Katz și colegii săi au fost pionieri în investigarea mecanismelor de transmitere sinaptică la joncțiunea neuromusculară. Ei au sugerat că canalul deschis de ACh era unul care avea permeabilitate egală atât cu Na+ cât și cu K+. Deoarece era la fel de permeabil la Na+ și K+, Katz a sugerat că, ca urmare a deschiderii acestor canale, potențialul membranei se va deplasa spre 0 MV. (O valoare a alfa în ecuația GHK egală cu una, care atunci când este înlocuită în ecuație, produce un potențial de aproximativ 0 MV.,)
figura 4.9
experimentul prezentat în figura din stânga testează acest concept. Celula musculară a fost penetrată cu un electrod de înregistrare, precum și cu un alt electrod care poate fi conectat la o sursă adecvată de potențial pentru a schimba artificial potențialul membranei. În mod normal, potențialul membranei este din nou de aproximativ -80 mV, se adaugă o cantitate mică de curare, astfel încât PPE să fie mic. Katz a observat în aceste experimente că mărimea PPE sa schimbat dramatic în funcție de potențialul celulei musculare., Dacă potențialul membranei este mutat la 0 mV, nu se înregistrează nicio modificare potențială. Dacă potențialul membranei este de + 30 mV, EPP este inversat. Deci, trei stimuli diferiți produc potențiale ale plăcii finale care sunt foarte diferite unele de altele.
lipsa unui răspuns atunci când potențialul este la 0 mV este deosebit de informativ. Luați în considerare motivul pentru care nu se înregistrează nicio schimbare potențială. Probabil, transmițătorul este eliberat și legat de receptori., Explicația simplă pentru o lipsă de schimbare potențială este că potențialul la care deschiderea canalelor ACh încearcă să ajungă a fost deja atins. Dacă potențialul membranei este mai pozitiv decât 0 mV, atunci EPP este inversat. Indiferent de potențial, schimbarea permeabilității tinde să deplaseze potențialul membranei spre 0 MV! Dacă potențialul de repaus este mai negativ decât 0 mV, există o deviere ascendentă. Dacă este mai pozitiv, există o deviere descendentă. Dacă este deja la 0 mV, nu există nici o deformare.
Figura 4.,10
acest potențial este numit și potențialul de inversare, deoarece este potențialul la care semnul potențialului sinaptic se inversează. Experimentul indică faptul că, ca urmare a legării ACh la receptori, canalele specifice devin la fel de permeabile la Na+ și K+. Această schimbare de permeabilitate tinde să deplaseze potențialul membranei de oriunde este inițial spre un nou potențial de 0 MV. de ce potențialul normal al plăcii de capăt nu atinge niciodată de fapt 0 MV?, Unul dintre motive este că secvența modificărilor de permeabilitate care stau la baza potențialului de acțiune „mlaștină” schimbările produse de PPE. Dar chiar dacă nu ar fi declanșat un potențial de acțiune, PPE încă nu ar atinge 0 mV. Acest lucru se datorează faptului că canalele ACh sunt doar o mică parte din numărul total de canale din fibrele musculare. Canalele K+ care înzestrează o celulă musculară cu potențialul său de repaus sunt de asemenea prezente. Treaba lor este să încerce să mențină celula la potențialul de odihnă., canalul deschis de ACh este un membru al unei clase generale de canale numite canale cu ligand sau receptori ionotropici. Așa cum este ilustrat în figura 4.10, site-ul de legare a emițătorului face parte din canalul în sine. Ca urmare a legării emițătorului de receptor (în general sunt necesare două molecule), există o schimbare conformațională a proteinei care permite deschiderea unei regiuni a porilor și curgerea ionilor în gradienții lor electrochimici. Detalii suplimentare ale canalului sunt prezentate în Capitolul 11.,
Testați-vă cunoștințele
- Întrebarea 1
- A
- B
- C
- D
un potențial endplate într-o celulă musculară scheletică ar putea fi, în principiu, produs de o permeabilitate scăzută la care dintre următorii ioni? (Să presupunem că există o permeabilitate inițială finită la fiecare dintre ionii enumerați mai jos și că gradienții de concentrație fiziologică sunt prezenți.):
A. Na+
B. Na+ și Ca2+
C. Ca2+
D., K+
un potențial endplate într-o celulă musculară scheletică ar putea fi, în principiu, produs de o permeabilitate scăzută la care dintre următorii ioni? (Să presupunem că există o permeabilitate inițială finită la fiecare dintre ionii enumerați mai jos și că gradienții de concentrație fiziologică sunt prezenți.):
A. Na+ acest răspuns este incorect.
un potențial al plăcii finale este o depolarizare care este produsă în mod normal prin creșterea simultană a permeabilității la ionii de sodiu și potasiu., Dacă a existat o scădere selectivă a permeabilității sodiului, o astfel de scădere nu ar duce la o depolarizare. Mai degrabă, ar duce de fapt la o hiperpolarizare, deoarece alfa în ecuația GHK ar fi redusă. Raportul permeabilității la sodiu și potasiu ar fi favorizat față de permeabilitatea potasiului, deplasând potențialul membranei spre potențialul de echilibru al potasiului și producând o hiperpolarizare.
B. Na+și Ca2+
C. Ca2 +
D., K+
un potențial endplate într-o celulă musculară scheletică ar putea fi, în principiu, produs de o permeabilitate scăzută la care dintre următorii ioni? (Să presupunem că există o permeabilitate inițială finită la fiecare dintre ionii enumerați mai jos și că gradienții de concentrație fiziologică sunt prezenți.):
A. Na+
B. Na+ și Ca2 + acest răspuns este incorect.
dacă permeabilitatea la sodiu și calciu ar fi scăzută, consecințele lor ar fi similare cu cele din alegerea A. scăderea permeabilității de sodiu în monoterapie ar tinde să hiperpolarizeze celula., În mod similar, scăderea permeabilității la calciu ar putea, de asemenea, să hiperpolarizeze celula. Potențialul de echilibru al calciului este o valoare foarte pozitivă, iar dacă ar exista o permeabilitate tonică de repaus la calciu, această permeabilitate ar contribui la o depolarizare tonică a potențialului membranei. Prin urmare, o scădere a permeabilității calciului ar elimina efectul depolarizant tonic și ar duce la o hiperpolarizare.
C. Ca2+
D., K+
un potențial endplate într-o celulă musculară scheletică ar putea fi, în principiu, produs de o permeabilitate scăzută la care dintre următorii ioni? (Să presupunem că există o permeabilitate inițială finită la fiecare dintre ionii enumerați mai jos și că gradienții de concentrație fiziologică sunt prezenți.):
A. Na+
B. Na+ și Ca2+
C. Ca2 + acest răspuns este incorect.
o scădere a permeabilității calciului singur ar produce, dacă ceva, o hiperpolarizare. Vezi logica răspunsului la alegerea B.
D., K+
un potențial endplate într-o celulă musculară scheletică ar putea fi, în principiu, produs de o permeabilitate scăzută la care dintre următorii ioni? (Să presupunem că există o permeabilitate inițială finită la fiecare dintre ionii enumerați mai jos și că gradienții de concentrație fiziologică sunt prezenți.):
A. Na+
B. Na+ și Ca2+
C. Ca2 +
D. K + acest răspuns este corect!
o scădere a permeabilității potasiului ar duce la o depolarizare similară cu un potențial de placă finală., Acest lucru se datorează faptului că există în repaus o permeabilitate tonică la potasiu și la sodiu. Permeabilitatea ridicată la potasiu tinde să mențină potențialul membranei în apropierea potențialului de echilibru al potasiului. Dacă această permeabilitate de repaus este scăzută, alfa în ecuația Goldman ar deveni o valoare mai mare, mutând potențialul membranei un pic mai aproape de potențialul de echilibru al sodiului (adică o depolarizare).