rysunek 4.1 (patrz powiększenie)
W związku z tym zrozumienie tej synapsy prowadzi do zrozumienia innych. Dlatego najpierw omówimy proces transmisji synaptycznej w szkieletowym węźle nerwowo-mięśniowym.,
cechy złącza synaptycznego przy złączu nerwowo-mięśniowym przedstawiono na rysunku po lewej stronie. Włókna mięśni szkieletowych są unerwiane przez neurony ruchowe, których ciała komórkowe znajdują się w rogu brzusznym rdzenia kręgowego. Region końcowy aksonu powoduje bardzo drobne procesy przebiegające wzdłuż komórek mięśni szkieletowych. Wzdłuż tych procesów są wyspecjalizowane struktury znane jako synapsy. Szczególną synapsę wykonywaną między neuronem ruchowym rdzenia a komórką mięśni szkieletowych nazywa się płytką końcową silnika ze względu na jego specyficzną strukturę.,
Synapsa na węźle nerwowo-mięśniowym ma trzy charakterystyczne cechy synaps chemicznych w układzie nerwowym. Po pierwsze, istnieje wyraźny rozdział między presynaptic i błony postsynaptic. Przestrzeń między nimi jest znana jako rozszczep synaptyczny. Przestrzeń mówi nam, że musi istnieć jakiś pośredni mechanizm sygnalizacyjny między neuronem presynaptycznym a neuronem postsynaptycznym, aby mieć przepływ informacji przez szczelinę synaptyczną. Po drugie, istnieje charakterystyczna wysoka gęstość małych pęcherzyków kulistych., Te pęcherzyki synaptyczne zawierają substancje neuroprzekaźnikowe. Synapsy są również związane z dużą gęstością mitochondriów. Po trzecie, w większości przypadków występuje charakterystyczne pogrubienie błony postsynaptycznej, co wynika przynajmniej częściowo z faktu, że błona postsynaptyczna ma wysoką gęstość wyspecjalizowanych receptorów, które wiążą chemiczne substancje nadajnikowe uwalniane z neuronu presynaptycznego. Dodatkowe szczegóły dotyczące cech morfologicznych połączeń synaptycznych przedstawiono w rozdziale 8 i rozdziale 10.
4.,2 fizjologia transmisji synaptycznej na węźle nerwowo-mięśniowym
rysunek 4.2
rysunek po prawej stronie przedstawia w bardzo schematyczny sposób, w jaki sposób możliwe jest dokładne zbadanie fizjologii transmisji synaptycznej na węźle nerwowo-mięśniowym szkieletowym. Kawałek mięśnia i dołączony do niego nerw umieszcza się w małej komorze Doświadczalnej wypełnionej odpowiednim roztworem Ringera. Potencjał spoczynkowy komórki mięśniowej jest rejestrowany za pomocą mikroelektrody. Elektrody są również umieszczone na powierzchni aksonu nerwu., Krótkie wstrząsy elektryczne powodują inicjowanie potencjałów akcji, które propagują się do terminala synaptycznego.
poniższy rysunek ilustruje dwa rodzaje potencjalnych zmian, które zostały zarejestrowane w takim izolowanym preparacie nerwowo-mięśniowym. Eksperyment ilustruje również właściwości potężnego leku, kurare, który okazał się bardzo przydatny w badaniu procesu transmisji synaptycznej w szkieletowym węźle nerwowo-mięśniowym. Część A ilustruje sekwencję potencjalnych zmian rejestrowanych w komórce mięśniowej w wyniku stymulowania aksonu motorycznego., Strzałka wskazuje punkt w czasie, w którym wstrząs jest dostarczany do osi silnika. Zauważ, że po wstrząsie jest okres spokoju. Opóźnienie wynika z czasu, jaki potrzeba, aby potencjał czynnościowy aksonu motorycznego rozprzestrzenił się z miejsca jego inicjacji. Po opóźnieniu w komórce mięśniowej rejestrowane są dwa rodzaje potencjałów. Po pierwsze, istnieje stosunkowo powoli zmieniający się potencjał, który będzie przedmiotem poniższej dyskusji., Jeśli ten wolny potencjał początkowy jest wystarczająco duży, jak zwykle w komórkach mięśni szkieletowych, w komórce mięśniowej pojawia się drugi potencjał, potencjał czynnościowy.
rysunek 4.3
potencjały działania w komórkach mięśni szkieletowych są spowodowane mechanizmami jonowymi podobnymi do omawianych wcześniej. W szczególności zachodzi zależna od napięcia zmiana przepuszczalności Na+, a następnie opóźniony wzrost przepuszczalności K+. (Dla komórek mięśni gładkich i mięśnia sercowego mechanizmy jonowe są jednak różne.,)
podstawowe zdarzenie, które wyzwala potencjał działania, można ujawnić, wykorzystując kurarę, truciznę używaną przez niektórych Indian Ameryki Południowej. Niska dawka kurary (część B) zmniejsza podstawowe zdarzenie, ale nadal nie jest wystarczająco zmniejszona, aby spaść poniżej progu. W przypadku podania nieco większej dawki kurary (część C), powolne Zdarzenie podstawowe staje się subthreshold. Sygnał bazowy jest znany jako potencjał płyty końcowej (EPP), ponieważ jest to zmiana potencjału rejestrowana na płycie końcowej silnika. Ogólnie rzecz biorąc, jest znany jako pobudzający potencjał postsynaptyczny (EPSP).,
Kurare blokuje potencjał płytki końcowej, ponieważ jest konkurencyjnym inhibitorem acetylocholiny (ACh), nadajnika uwalnianego w terminalu presynaptycznym. Curare nie blokuje zależnej od napięcia na + konduktancji ani zależnej od napięcia K+ konduktancji, która stanowi podstawę potencjału czynnościowego mięśni. Kurara wpływa na bodziec (EPSP), który zwykle prowadzi do inicjacji potencjału czynnościowego mięśni. Zwierzę zatrute kurarą udusi się, ponieważ proces transmisji nerwowo-mięśniowej w mięśniach oddechowych jest zablokowany.,
zwykle wielkość potencjału płyty końcowej jest dość duża. Rzeczywiście, Amplituda potencjału płyty końcowej wynosi około 50 mV, ale tylko około 30 mV jest potrzebne do osiągnięcia progu. Dodatkowe 20 mV nazywa się współczynnikiem bezpieczeństwa. 40 mV w amplitudzie) z powodu zmęczenia, EPP osiągnie próg i zachowana zostanie relacja jeden do jednego między potencjałem czynnościowym w aksonie motorycznym a potencjałem czynnościowym w komórce mięśniowej.
rysunek 4.4
4.,3 propagacja PPE
Jakie są właściwości PPE i jak porównuje się z właściwościami potencjału działania?
czy potencjał krańcowy jest spowodowany zależną od napięcia zmianą przenikalności Na+ I K+, np. potencjałem działania?
czy potencjał endplate propagowany jest w sposób „wszystko albo nic” jak potencjał działania?
rysunek po lewej stronie ilustruje eksperyment, który bada propagację potencjału płyty końcowej. Włókno mięśniowe jest wielokrotnie nabijane elektrodami w odstępach 1 mm., (Zauważ, że potencjał płyty końcowej jest mały, ponieważ eksperyment ten odbywa się w obecności niskiego stężenia kurary, więc potencjał płyty końcowej może być rejestrowany bez komplikacji związanych z wyzwalaniem potencjału czynnościowego.) Potencjał płyty końcowej nie jest propagowany w sposób „wszystko albo nic”. Rozprzestrzenia się wzdłuż mięśni, ale robi to z zanikiem., Tak więc, rozprzestrzenianie się potencjału płyty końcowej z miejsca inicjacji do innych miejsc wzdłuż komórki mięśniowej następuje biernie i ze zmniejszeniem, podobnie jak zmiana potencjału subthreshold w jednej części aksonu rozprzestrzenia się wzdłuż aksonu, lub po prostu jak zmiana temperatury w jednym punkcie na metalowym pręta rozprzestrzenia się wzdłuż pręta.
4.4 przegląd sekwencji zdarzeń leżących u podstaw EPP
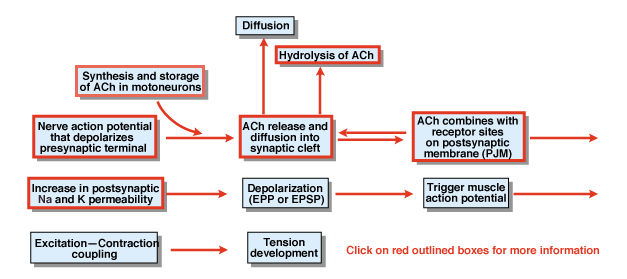
rysunek 4.5
Jakie są inne etapy procesu chemicznej transmisji synaptycznej? Rysunek 4.,5 zawiera przegląd. Potencjał działania nerwów, który jest inicjowany w ciele komórkowym neuronu ruchowego rdzenia rozprzestrzenia się korzeni brzusznych i ostatecznie atakuje terminale synaptyczne neuronów ruchowych. W wyniku potencjału czynnościowego acetylocholina transmitera chemicznego (ACh) jest uwalniana do szczeliny synaptycznej. ACh dyfunduje przez rozszczep synaptyczny i wiąże się ze specjalnymi receptorami na błonie postsynaptycznej lub postjunkcyjnej. Wiązanie ACh z receptorami powoduje zmianę konformacyjną w kanale błonowym, który jest szczególnie przepuszczalny zarówno dla Na+, jak i K+., W wyniku wzrostu przepuszczalności Na + I K+ następuje depolaryzacja błony postsynaptycznej. Ta depolaryzacja nazywana jest potencjałem endplate lub bardziej ogólnie EPSP. Jeśli EPSP jest wystarczająco duży, jak zwykle znajduje się na złączu nerwowo-mięśniowym, prowadzi to do inicjacji potencjału czynnościowego w komórce mięśniowej. Potencjał działania inicjuje proces sprzężenia skurczowego wzbudzenia i rozwój napięcia. Czas trwania potencjału płyty końcowej wynosi około 10 msec.
dwa czynniki kontrolują czas trwania EPSP na węźle nerwowo-mięśniowym., Po pierwsze, ACh jest usuwany przez dyfuzję. Po drugie, substancja w rozszczepieniu synaptycznym, zwana acetylocholinoesterazą (AChE), hydrolizuje lub rozkłada ACh. Ból jest jednym z najbardziej wydajnych enzymów znanych. Pojedyncza cząsteczka AChE może hydrolizować 600 000 cząsteczek ACh na minutę.
4, 5 rola bólu
Rysunek 4, 6
ważna rodzina substancji, z których jedną jest Neostygmina, hamuje działanie bólu. Neostygmina blokuje działanie bólu, a tym samym zwiększa potencjał płytki końcowej i wydłuża jej czas trwania., Rysunek ten ilustruje dwa potencjały krańcowe. Jedną zarejestrowano w soli fizjologicznej i kurarze, a drugą po dodaniu neostygminy do roztworu. (Kurare dodaje się tak, że właściwości EPP mogą być badane bez wyzwalania potencjału czynnościowego w komórce mięśniowej.) Po zastosowaniu neostygminy potencjał płytki końcowej jest znacznie większy i dłuższy.
4.6 A miastenia
miastenia jest związana z ciężkim osłabieniem mięśni z powodu zmniejszenia liczby receptorów acetylocholiny w komórce mięśniowej., Jeśli potencjał płyty końcowej jest mniejszy, potencjał płyty końcowej nie osiągnie progu. Jeśli nie osiągnie progu, nie będzie żadnego potencjału czynnościowego w komórce mięśniowej i nie będzie skurczu mięśnia, co powoduje osłabienie mięśni. Neostygmina i inne inhibitory AChE są stosowane w leczeniu pacjentów z miastenią. Środki te sprawiają, że ilość acetylocholiny, która jest uwalniana, skuteczniej dociera do pozostałych receptorów acetylocholiny.
4.,6b środki nerwowe
chociaż inhibitory AChE mają ważną wartość terapeutyczną, niektóre inhibitory były i są nadal stosowane jako trucizny. Niektóre inhibitory AChE, takie jak Soman i Sarin tworzą dość nieodwracalny blok bólu. Blok ten prowadzi do ekstremalnych poziomów ACh w szczelinie synaptycznej. Osoby tak zatrute umierają z powodu drgawek i spastyczności mięśni, w tym mięśni oddechowych.
rysunek 4.7
4.,7 jontoforeza ACh
jontoforeza jest interesującą techniką, która może być wykorzystana do dalszego badania hipotezy, że ACh jest substancją neuroprzekaźnika w węźle nerwowo-mięśniowym. Jeśli ACh jest nadajnikiem, który jest uwalniany przez tę synapsę, można by przewidzieć, że powinno być możliwe zastąpienie sztucznego zastosowania nadajnika normalnym zwolnieniem nadajnika. Ponieważ ACh jest dodatnio naładowaną cząsteczką, może być wymuszona z mikroelektrody w celu symulacji uwalniania ACh z terminala presynaptycznego.
Rysunek 4.,8
Rysunek 4.8 porównuje EPP wytwarzany przez stymulację aksonu motorycznego i reakcję na wyrzuty ACh. Potencjalna zmiana wygląda prawie identycznie jak potencjał płyty końcowej wytwarzany przez normalne uwalnianie ACh. Eksperyment ten zapewnia eksperymentalne wsparcie dla koncepcji, że ACh jest naturalnym nadajnikiem na tej synapsie.,
odpowiedź na wyrzut ACh ma kilka innych ciekawych właściwości, które są zgodne z cholinergiczną naturą synapsy w szkieletowym węźle nerwowo-mięśniowym. Neostygmina sprawia, że reakcja na jontoforezę ACh jest dłuższa i większa. Kurare zmniejsza odpowiedź, ponieważ konkuruje z normalnym wiązaniem ACh. Jeśli ACh jest wyrzucany do komórki mięśniowej, nic się nie dzieje, ponieważ receptory acetylocholiny nie znajdują się wewnątrz; są one na zewnątrz komórki mięśniowej., Zastosowanie acetylocholiny do regionów mięśni z dala od płytki końcowej nie powoduje odpowiedzi, ponieważ receptory ACh są skoncentrowane w regionie synaptycznym.
aby sprawdzić swoje dotychczasowe zrozumienie, zastanów się, w jaki sposób środek taki jak TTX wpłynie na generowanie zarówno EPP, jak i odpowiedź włókna mięśniowego na jontoforetyczne zastosowanie ACh? TTX nie ma wpływu na odpowiedź na ACh, ale blokuje EPP. Powód, dla którego odpowiedź na ACh jest nienaruszona, jest jasny, ale wielu oczekuje, że jeśli nie ma żadnego efektu w tym przypadku, nie powinno mieć to również wpływu na EPL., Tetrodotoksyna nie wpływa na Wiązanie acetylocholiny z receptorami, a zatem nie wpływa na odpowiedź na bezpośrednie zastosowanie ACh. Jednak tetrodotoksyna wpływa na zdolność wywoływania potencjału czynnościowego w aksonie motorycznym. Jeśli w aksonie silnika nie można wywołać potencjału działania, nie może to spowodować uwolnienia nadajnika. W ten sposób tetrodotoksyna całkowicie zlikwidowałaby EPL. Blok nie byłby spowodowany blokiem receptorów ACh, ale raczej blokiem jakiegoś kroku przed uwolnieniem nadajnika.
4.,8 mechanizmy jonowe EPP
Bernard Katz i jego współpracownicy byli pionierami w badaniu mechanizmów transmisji synaptycznej na węźle nerwowo-mięśniowym. Zasugerowali, że kanał otwarty przez ACh jest taki, który ma równą przepuszczalność zarówno dla Na+ , jak i K+. Ponieważ była ona równie przepuszczalna dla Na+ I K+, Katz zasugerował, że w wyniku otwarcia tych kanałów potencjał membrany przesunie się w kierunku 0 mV. (Wartość alfa w równaniu GHK równa Jedynce, która po podstawianiu do równania daje potencjał około 0 mV.,)
rysunek 4.9
eksperyment pokazany na rysunku po lewej stronie testuje tę koncepcję. Komórka mięśniowa została penetrowana elektrodą rejestrującą, a także inną elektrodą, którą można podłączyć do odpowiedniego źródła potencjału w celu sztucznej zmiany potencjału błonowego. Zwykle potencjał membrany wynosi około -80 mV, dodaje się niewielką ilość kurary, dzięki czemu EPP jest mały. Katz zauważył w tych eksperymentach, że rozmiar EPP zmienił się dramatycznie w zależności od potencjału komórki mięśniowej., Jeśli potencjał membranowy zostanie przesunięty do 0 mV, nie zostanie zarejestrowana żadna zmiana potencjału. Jeśli potencjał membrany wynosi +30 mV, EPP jest odwrócony. Tak więc trzy różne bodźce wytwarzają potencjały endplate, które są bardzo różne od siebie.
Brak odpowiedzi, gdy potencjał wynosi 0 mV jest szczególnie pouczający. Zastanów się, dlaczego nie zarejestrowano żadnej potencjalnej zmiany. Prawdopodobnie Nadajnik jest uwalniany i wiąże się z receptorami., Prostym wyjaśnieniem braku potencjalnej zmiany jest to, że potencjał, do którego próbuje się dotrzeć otwarcie kanałów ACh, został już osiągnięty. Jeśli potencjał membrany jest bardziej dodatni niż 0 mV, to EPP jest odwrócony. Bez względu na potencjał, zmiana przepuszczalności ma tendencję do przesuwania potencjału membrany w kierunku 0 mV! Jeśli potencjał spoczynkowy jest bardziej ujemny niż 0 mV, występuje odchylenie w górę. Jeśli jest bardziej pozytywny, istnieje odchylenie w dół. Jeśli jest już przy 0 mV, nie ma ugięcia.
Rysunek 4.,10
potencjał ten nazywany jest również potencjałem odwrotnym, ponieważ jest to potencjał, przy którym znak potencjału synaptycznego ulega odwróceniu. Eksperyment wskazuje, że w wyniku wiązania ACh z receptorami określone kanały stają się równie przepuszczalne dla Na+ I K+. Ta zmiana przepuszczalności ma tendencję do przenoszenia potencjału membranowego z dowolnego miejsca, gdzie jest początkowo, w kierunku nowego potencjału 0 mV.
Dlaczego normalny potencjał krańcowy nigdy nie osiąga 0 mV?, Jednym z powodów jest to, że sekwencja zmian przepuszczalności, które leżą u podstaw potencjału działania, „wypiera” zmiany wytwarzane przez EPL. Ale nawet gdyby nie został uruchomiony potencjał działania, EPL nadal nie osiągnęłaby 0 mV. Dzieje się tak dlatego, że kanały ACh stanowią tylko niewielki ułamek całkowitej liczby kanałów we włóknach mięśniowych. Obecne są również kanały K+, które nadają komórce mięśniowej jej potencjał spoczynkowy. Ich zadaniem jest utrzymanie komórki w stanie spoczynku.,
kanał otwarty przez ACh należy do ogólnej klasy kanałów zwanych kanałami bramkowanymi ligandami lub receptorami jonotropowymi. Jak pokazano na rysunku 4.10, miejsce wiązania nadajnika jest częścią samego kanału. W wyniku wiązania nadajnika z receptorem (na ogół konieczne są dwie cząsteczki) dochodzi do zmiany konformacyjnej w białku, która umożliwia otwarcie obszaru porów i przepływ jonów w dół ich gradientów elektrochemicznych. Dodatkowe szczegóły dotyczące kanału przedstawiono w rozdziale 11.,
Sprawdź swoją wiedzę
- Pytanie 1
- a
- B
- C
- D
potencjał płytki końcowej w komórce mięśni szkieletowych może w zasadzie być wytwarzany przez zmniejszoną przepuszczalność, do którego z następujących jonów? (Załóżmy, że istnieje skończona wstępna przepuszczalność dla każdego z wymienionych poniżej jonów i że występują gradienty stężenia fizjologicznego.):
A. Na +
B. Na+i Ca2 +
C. Ca2+
D., K +
potencjał endplate w komórce mięśni szkieletowych może w zasadzie być wytwarzany przez zmniejszoną przepuszczalność, do którego z następujących jonów(y)? (Załóżmy, że istnieje skończona wstępna przepuszczalność dla każdego z wymienionych poniżej jonów i że występują gradienty stężenia fizjologicznego.):
A. Na+ ta odpowiedź jest niepoprawna.
potencjał płytki końcowej to depolaryzacja, która jest zwykle wytwarzana przez jednoczesne zwiększenie przepuszczalności jonów sodu i potasu., Jeśli wystąpi selektywne zmniejszenie przepuszczalności sodu, takie zmniejszenie nie prowadziłoby do depolaryzacji. Raczej prowadziłoby to do hiperpolaryzacji, ponieważ alfa w równaniu GHK zostałaby zmniejszona. Stosunek przepuszczalności do sodu i potasu byłby faworyzowany w kierunku przepuszczalności potasu, przenosząc potencjał membranowy w kierunku potencjału równowagi potasowej i wytwarzając hiperpolaryzację.
B. Na+i Ca2 +
C. Ca2+
D., K +
potencjał endplate w komórce mięśni szkieletowych może w zasadzie być wytwarzany przez zmniejszoną przepuszczalność, do którego z następujących jonów(y)? (Załóżmy, że istnieje skończona wstępna przepuszczalność dla każdego z wymienionych poniżej jonów i że występują gradienty stężenia fizjologicznego.):
A. Na+
B. Na+ i Ca2+ ta odpowiedź jest niepoprawna.
gdyby zmniejszono przepuszczalność sodu i wapnia, ich konsekwencje byłyby podobne do tych z wyboru A. samo zmniejszenie przepuszczalności sodu prowadziłoby do hiperpolaryzacji komórki., Podobnie, zmniejszenie przepuszczalności wapnia może również hiperpolaryzować komórkę. Potencjał równowagi wapnia jest bardzo pozytywną wartością, a jeśli istnieje pewna tonizująca przepuszczalność spoczynkowa wapnia, przepuszczalność ta przyczyniłaby się do tonicznej depolaryzacji potencjału błony. Stąd spadek przepuszczalności wapnia usunąłby ten toniczny efekt depolaryzacji i spowodowałby hiperpolaryzację.
C. Ca2+
D., K +
potencjał endplate w komórce mięśni szkieletowych może w zasadzie być wytwarzany przez zmniejszoną przepuszczalność, do którego z następujących jonów(y)? (Załóżmy, że istnieje skończona wstępna przepuszczalność dla każdego z wymienionych poniżej jonów i że występują gradienty stężenia fizjologicznego.):
A. Na+
B. Na+ i Ca2+
C. Ca2+ ta odpowiedź jest nieprawidłowa.
samo zmniejszenie przepuszczalności wapnia spowodowałoby hiperpolaryzację. Patrz logika odpowiedzi na wybór B.
D., K +
potencjał endplate w komórce mięśni szkieletowych może w zasadzie być wytwarzany przez zmniejszoną przepuszczalność, do którego z następujących jonów(y)? (Załóżmy, że istnieje skończona wstępna przepuszczalność dla każdego z wymienionych poniżej jonów i że występują gradienty stężenia fizjologicznego.):
A. Na+
B. Na+ i Ca2+
C. Ca2+
D. K+ ta odpowiedź jest prawidłowa!
zmniejszenie przepuszczalności potasu prowadziłoby do depolaryzacji podobnej do potencjału płytki końcowej., Dzieje się tak dlatego, że w spoczynku występuje toniczna przepuszczalność potasu i sodu. Wysoka przepuszczalność potasu ma tendencję do utrzymywania potencjału membranowego w pobliżu potencjału równowagi potasowej. Jeśli ta przepuszczalność spoczynkowa zostanie zmniejszona, alfa w równaniu Goldmana stanie się większą wartością, przesuwając potencjał błonowy nieco bliżej potencjału równowagi sodowej (tj. depolaryzacji).