wprowadzenie
wielki wgląd Darwina i jednocząca zasada biologii dzisiaj, jest taka, że wszystkie gatunki są spokrewnione ze sobą jak siostry, kuzynki i dalekie krewne w rozległym drzewie genealogicznym życia. Implikacje są zapierające dech w piersiach; gdybyśmy mogli cofnąć się wystarczająco daleko w czasie, znaleźlibyśmy wspólnych przodków między sobą i każdym innym żywym organizmem, od jeżozwierzy, przez flamingi, po Kaktusy., Nasza najbliższa rodzina ewolucyjna składa się z małp człekokształtnych, grupy naczelnych, która obejmuje „małpy mniejsze” (syjamangi i gibony), jak również „wielkie małpy” (szympansy, bonobos, goryle i orangutany). Wśród wielkich małp, naszymi najbliższymi krewnymi są szympansy i bonobos (ryc. Zapis kopalny wraz z badaniami ludzkiego i Małpiego DNA wskazuje, że ludzie mieli wspólnego przodka z szympansami i bonobosami około 6 milionów lat temu (mya)., Zaczynamy tę dyskusję na temat ewolucji naszego gatunku w Afryce, pod koniec geologicznego okresu znanego jako miocen, tuż przed tym, jak nasze rodowody oddzieliły się od szympansów i bonobosów.
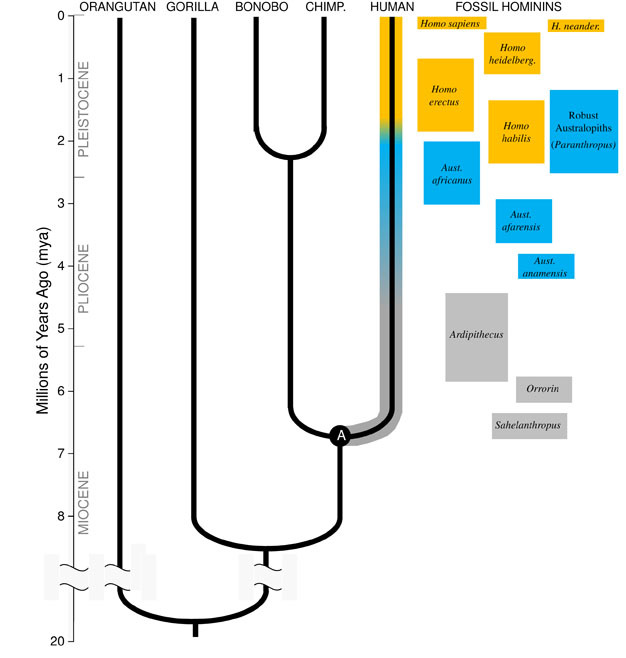
mioceńskie pochodzenie rodu Homininów
aby zrozumieć ewolucję jakiegokolwiek gatunku, musimy najpierw ustalić jego stan przodków: z jakiego rodzaju zwierzęcia wyewoluował? Dla naszej linii wymaga to, abyśmy spróbowali zrekonstruować ostatniego wspólnego przodka ludzi i szympansów (oznaczonego „A” na rysunku 1). Ostatni wspólny przodek człowieka-szympansa (HC-LCA) to gatunek, od którego oddzieliła się linia homininów i szympansów & linia bonobo., Hominidy są gatunkami na naszej gałęzi drzewa małp człekokształtnych po podziale z szympansem& linii bonobo, obejmującej wszystkie wymarłe gatunki i ewolucyjne gałęzie boczne (ryc. 1).
w miocenie stwierdzono dużą różnorodność gatunków małp, z dziesiątkami gatunków znanych z zapisu kopalnego w Afryce, Europie i Azji. Gatunki te różnią się anatomią i ekologią i nie jest jasne, który z odkrytych dotąd gatunków kopalnych reprezentuje HC-LCA (Kunimatsu et al. 2007; Young and MacLatchy, 2004)., Niemniej jednak, wiemy z skamieniałości i dowodów porównawczych, że był on znacznie bardziej podobny do żywych małp niż do żywych ludzi. HC-LCA miał mózg i ciało wielkości małpy, ze stosunkowo długimi rękami i palcami oraz chwytną stopą, która pozwalała mu żerować na drzewach. Zęby psów były prawdopodobnie duże i ostre, jak u kilku mioceńskich małp człekokształtnych. Co więcej, kły były prawdopodobnie dymorficzne płciowo, a samce miały znacznie większe kły niż samice, co widać u żyjących małp i skamieniałości z miocenu., Podobnie jak żyjące małpy, chodził po ziemi czworonożnie (na czworakach), a jego dieta składała się prawie wyłącznie z pokarmu roślinnego, głównie owoców i liści.
wczesne hominidy
zmiany anatomii małpopodobnych są zauważalne w skamieniałościach małp człekokształtnych z późnego miocenu w Afryce. Niektóre gatunki małp człekokształtnych z tego okresu wykazują cechy typowe dla ludzi, ale nie są widoczne u innych żyjących małp człekokształtnych, co prowadzi paleoantropologów do wniosku, że te skamieniałości reprezentują wczesnych członków linii homininów., Pierwsze ludzkie cechy, które pojawiły się w zapisie kopalnym homininów, to dwunożne chodzenie i mniejsze, tępe kły.
najstarszymi homininami obecnie znanymi są Sahelanthropus tchadensis z Czadu (Brunet et al. 2005) i Orrorin tugenensis z Kenii (Senut et al. 2001). Sahelanthropus, datowany na 6-7 milionów lat, jest znany z w dużej mierze kompletnej czaszki i kilku innych fragmentarycznych szczątków. Jego rozmiar mózgu, 360cc, mieści się w zakresie obserwowanym u szympansów, a czaszka ma masywny grzbiet brwiowy, podobny do grubości samców goryli (Brunet et al. 2005)., Jednak położenie i orientacja otworu magnum, otworu w podstawie czaszki, przez który przechodzi rdzeń kręgowy, sugeruje, że Sahelanthropus stał i chodził dwunożnie, z kręgosłupem trzymanym pionowo, jak u współczesnych ludzi, a nie poziomo, jak u małp i innych czworonogów (Zollikofer et al. 2005). Orrorin znany jest przede wszystkim ze skamieniałości pozaczaszkowych, w tym częściowej kości udowej. Proksymalna część kości udowej wykazuje podobieństwa do tych współczesnych ludzi, co sugeruje, że gatunek był dwunożny (Pickford et al. 2002)., Nie odnaleziono czaszek Orrorina, więc morfologia czaszki i wielkość mózgu są niepewne. Zarówno u Orrorina, jak i Sahelanthropusa zęby psów samców są większe i bardziej spiczaste niż u współczesnych ludzi, ale są małe i tępe w porównaniu do psów małp człekokształtnych. Sugeruje to, że dymorfizm płciowy psów — a co za tym idzie, konkurencja między samcami o dostęp do godów i samic — była zmniejszona u tych wczesnych hominidów w porównaniu z wielkimi małpami.
zdecydowanie najbardziej znanym wczesnym homininem jest Ardipithecus ramidus, 4.,4 mln lat gatunków z Etiopii, który jest znany z prawie kompletny szkielet, jak również liczne inne szczątki zębów i szkieletu (White et al. 2009). Ar. ramidus i starszy, spokrewniony z nim gatunek znany z fragmentarycznych szczątków, Ar. kadabba (5,8-5,2 mya), mają zredukowane kły podobne do tych z Orrorin i Sahelanthropus. Czaszka Ar. ramidus jest raczej małpopodobny i ogólnie podobny do Sahelanthropusa, z małym mózgiem szympansa wielkości 300 – 350cc (ryc. 2). Szkielet pozaczaszkowy Ardipithecus jest intrygujący., Chociaż odnaleziona miednica jest bardzo rozdrobniona, wykazuje morfologię zupełnie inną niż u żyjących małp, z krótszym, bardziej przypominającym miskę kształtem, który sugeruje, że Ardipithecus chodził dwunożnie; jest to zgodne z pozycją otworu magnum, co sugeruje wyprostowaną postawę. Jednak jego długie kończyny przednie i palce oraz rozbieżny, chwytny pierwszy palec (hallux) sugerują, że Ardipithecus spędzał większość czasu na drzewach. Ogólne wrażenie jest w dużej mierze nadrzewnym gatunkiem, który chodził dwunożnie, gdy tylko odważył się na ziemię.,
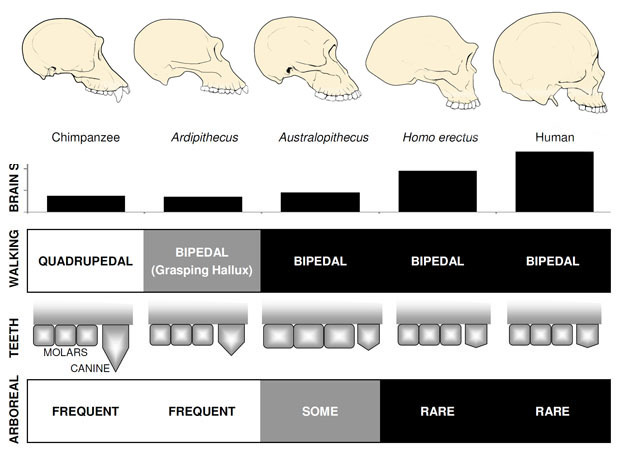
Australopithecus
około 4mya znajdujemy najwcześniejszych przedstawicieli rodzaju Australopithecus, homininów, które były biegłymi dwunożnymi, ale nadal używali drzew do jedzenia i ochrony. Pierwsze okazy Australopithecus odkryto w Afryce Południowej w 1924 roku (Dart, 1925), a wysiłki badawcze w ciągu kolejnych ośmiu dekad przyniosły setki skamieniałości z kilku gatunków w całej Afryce Wschodniej i Południowej., Obecnie wiemy, że Australopithecus był bardzo udanym rodzajem, który przetrwał prawie trzy miliony lat (Rysunek 1).
najbardziej znanymi gatunkami Australopithecus są A. afarensis (3,6-2,9 mya) z Afryki Wschodniej i A. africanus (3,2–2,0 mya) z Afryki Południowej., Miednica i kończyna dolna tych gatunków wyraźnie wskazuje, że były w pełni dwunożne: miednica jest krótka i w kształcie miski, doprowadzając mięśnie pośladkowe do boku ciała, jak u współczesnych ludzi, do stabilizacji tułowia podczas dwunożności, a pierwszy palec jest zgodny z innymi palcami (Ward, 2002; Harcourt-Smith and Aiello, 2004). Stopa Australopithecus mogła mieć nawet łuk podobny do człowieka, na podstawie analizy śródstopia i skamieniałych stóp Laetoli (Ward et al. 2011)., Niemniej jednak, w porównaniu do współczesnych ludzi, przedramiona były długie, a palce u rąk i nóg były długie i nieco zakrzywione, co sugeruje, że Australopithecus regularnie używał drzew do żerowania i być może jako schronienia przed drapieżnikami w nocy. Ta mieszana naziemna& strategia arborealna dobrze służyłaby tym gatunkom w mieszanych środowiskach leśnych i sawannowych, które zamieszkiwały.
rozmiar mózgu u Australopithecus wahał się od 390 do 515cc, podobny do szympansów i goryli (Falk et al., 2000), sugerując, że zdolności poznawcze były zasadniczo podobne do żyjących małp (ryc. 2). Rozmiar ciała u Australopithecus był raczej mały i dymorficzny płciowo, około 30 kg dla samic i 40 kg dla samców (Mchenry, 1992). Ten poziom dymorfizmu nie znajduje odzwierciedlenia w kłach, które były małe, tępe i monomorficzne, jak u wcześniejszych homininów.
w przeciwieństwie do kłów zęby trzonowe u australopiteków były znacznie większe niż u wcześniejszych hominidów i miały grubsze szkliwo. Sugeruje to, że ich dieta obejmowała twarde, niskiej jakości pokarmy roślinne, które wymagały silnego żucia do przetworzenia., Podgrupa Australopithecus, znana jako” solidne ” Australopithecus (często określana jako osobny rodzaj Paranthropus) z powodu ich ogromnych zębów i mięśni żujących, przyjęła tę adaptację do skrajności. Większość gatunków Australopithecus wymarła o 2 mln lat, ale niektóre solidne formy przetrwały do około 1,2 mln lat we wschodniej i Południowej Afryce.
rodzaj Homo
najwcześniejsze skamieniałości naszego rodzaju, Homo, znajdują się w Afryce Wschodniej i datowane na 2,3 mya (Kimbel et al. 1997)., Te wczesne okazy są podobne pod względem wielkości mózgu i ciała do Australopithecus, ale wykazują różnice w zębach trzonowych, co sugeruje zmianę diety. W rzeczy samej, o co najmniej 1,8 mln lat wczesni członkowie naszego rodzaju używali prymitywnych kamiennych narzędzi do rzeźbienia zwłok zwierzęcych, dodając bogate w energię mięso i szpik kostny do diety opartej na roślinach.
najstarszy członek rodzaju Homo, H. habilis (2,3–1,4 mya) znajduje się w Afryce Wschodniej i jest związany z zarżniętych kości zwierząt i prostych narzędzi kamiennych (Blumenschine et al. 2003). Jego bardziej groźny i rozpowszechniony potomek, H., erectus, występuje w całej Afryce i Eurazji i utrzymuje się od 1,9 mya do 100 kya, a być może nawet później (Anton, 2003). Podobnie jak współcześni ludzie, H. erectus nie posiadał adaptacji kończyn przednich do wspinaczki widzianej u Australopithecus(ryc. 2). Jego globalna ekspansja sugeruje, że H. erectus był ekologicznie elastyczny, z zdolnością poznawczą do adaptacji i rozwoju w bardzo różnych środowiskach. Nic dziwnego, że to właśnie u H. erectus zaczynamy zauważać znaczny wzrost wielkości mózgu, do 1250 cm3 dla późniejszych okazów azjatyckich (Anton, 2003). Wielkość molowa jest zmniejszona w H., erectus w stosunku do Australopithecus, odzwierciedlając jego łagodniejszą, bogatszą dietę.
Około 700 kya, a być może wcześniej, H. erectus w Afryce dał początek H. heidelbergensis, gatunkowi bardzo podobnemu do nas pod względem proporcji ciała, adaptacji zębów i zdolności poznawczych (Rightmire, 2009). H. heidelbergensis, często określany jako” archaiczny ” Homo sapiens, był aktywnym łowcą dużych zwierząt, produkował zaawansowane narzędzia w stylu Levallois i przez co najmniej 400 kya nauczył się kontrolować ogień (Roebroeks and Villa, 2011). Neandertalczycy (H., neanderthalensis), zaadaptowane na zimno homininy o grubej budowie ciała, złożonych zachowaniach i mózgach podobnych do naszego, uważa się, że wyewoluowały z populacji H. heidelbergensis w Europie o co najmniej 250 kya (Rightmire, 2008; Hublin, 2009).
dowody kopalne i DNA sugerują, że nasz własny gatunek, H. sapiens, wyewoluował w Afryce 200 kya (Relethford, 2008; Rightmire, 2009), prawdopodobnie od H. heidelbergensis. Wzrost wyrafinowania behawioralnego H., sapiens, na co wskazują nasze duże mózgi (1400 cm3) i archeologiczne dowody szerszego zestawu narzędzi i sprytnych technik łowieckich, pozwoliły naszemu gatunkowi rozkwitnąć i rosnąć na kontynencie afrykańskim. Do 100kya nasz gatunek rozlał się do Eurazji, ostatecznie rozszerzając się na całym świecie do Australii i Obu Ameryk (DiGiorgio et al. 2009). Po drodze nasz gatunek wyparł inne napotkane hominidy, w tym neandertalczyków w Europie i podobne formy w Azji. (Zauważ, że nie wszyscy zgadzają się z tą interpretacją danych, patrz Tryon i Bailey)., Badania starożytnego DNA ekstrahowanego ze skamieniałości neandertalczyków sugerują, że nasz gatunek mógł od czasu do czasu krzyżować się z nimi (Green et al., 2010). Nasz rosnący globalny wpływ trwa nadal, ponieważ innowacje kulturowe, takie jak rolnictwo i urbanizacja, kształtują krajobraz i gatunki wokół nas.
podsumowanie
ewolucja naszego gatunku od małpopodobnego przodka miocenu była złożonym procesem. Nasza linia jest pełna bocznych gałęzi i ewolucyjnych ślepych zaułków, z takimi gatunkami jak wytrzymałe australopithmy, które przetrwały ponad milion lat, zanim zanikły., Niektóre cechy ludzkie, takie jak dwunożność, ewoluowały bardzo wcześnie, podczas gdy inne, takie jak duże mózgi, nie ewoluowały aż do stosunkowo niedawna. Jeszcze inne cechy, takie jak wielkość trzonu, ewoluowały w jednym kierunku, aby później zostać wyparte przez zmieniające się naciski ekologiczne. Zamiast potężnego statku wyznaczającego prosty kurs w kierunku określonego z góry celu, ewolucja naszej linii-w rzeczy samej, linii jakiegokolwiek gatunku-pasuje do obrazu łodzi ratunkowej rzucanej przez zmieniające się Morza zmian środowiska, genetycznego szczęścia i geologicznego przypadku. Zastanawia się, dokąd nas zaprowadzi następne sześć milionów lat.