figuur 4.1 (zie vergrote weergave)
bijgevolg leidt een begrip van deze synapse tot een begrip van de anderen. Daarom zullen we eerst het proces van synaptische transmissie op de skeletale neuromusculaire junctie bespreken.,
De kenmerken van de synaptische kruising bij de neuromusculaire kruising worden weergegeven in de figuur links. Skeletspiervezels worden innervated door motorneuronen waarvan de cellichamen zich in de ventrale hoorn van het ruggenmerg bevinden. Het terminale gebied van het axon geeft aanleiding tot zeer fijne processen die langs skeletspiercellen lopen. Langs deze processen zijn gespecialiseerde structuren bekend als synapsen. De specifieke synaps gemaakt tussen een spinale Motor neuron en skeletspiercel wordt de motor endplate genoemd vanwege zijn specifieke structuur.,
de synaps op de neuromusculaire kruising heeft drie karakteristieke kenmerken van chemische synapsen in het zenuwstelsel. Eerst, is er een duidelijke scheiding tussen presynaptic en het postsynaptic membraan. De ruimte tussen de twee staat bekend als de synaptische spleet. De ruimte vertelt ons dat er een intermediair signaleringsmechanisme moet zijn tussen het presynaptische neuron en het postsynaptische neuron om informatie door de synaptische spleet te laten stromen. Ten tweede is er een karakteristieke hoge dichtheid van kleine bolvormige blaasjes., Deze synaptische blaasjes bevatten neurotransmitterstoffen. Synapsen worden ook geassocieerd met een hoge dichtheid van mitochondria. Ten derde is er in de meeste gevallen een kenmerkende verdikking van het postsynaptisch membraan, wat ten minste gedeeltelijk te wijten is aan het feit dat het postsynaptisch membraan een hoge dichtheid van gespecialiseerde receptoren heeft die de chemische zenderstoffen binden die vrijkomen uit het presynaptisch neuron. Aanvullende details over de morfologische kenmerken van synaptische kruisingen worden gegeven in hoofdstuk 8 en hoofdstuk 10.
4.,2 Physiology of Synaptic Transmission at the Neuromuscular Junction
figuur 4.2
de figuur rechts illustreert op een zeer schematische manier hoe het mogelijk is om de fysiologie van synaptic transmission at the skeletal neuromuscular junction gedetailleerd te bestuderen. Een stuk spier en de aangehechte zenuw worden geplaatst in een kleine experimentele kamer gevuld met een geschikte Ringer oplossing. Het rustpotentieel van de spiercel wordt geregistreerd met een micro-elektrode. Elektroden worden ook geplaatst op het oppervlak van het zenuwaxon., Korte elektrische schokken veroorzaken actiepotentialen te worden geïnitieerd, die zich verspreiden naar de synaptische terminal.
onderstaande figuur illustreert twee soorten potentiële veranderingen die werden geregistreerd in zo ‘ n geïsoleerde zenuwspiervoorbereiding. Het experiment illustreert ook de eigenschappen van een krachtig medicijn, curare, dat zeer nuttig is gebleken bij het bestuderen van het proces van synaptische transmissie bij de neuromusculaire kruising van het skelet. Deel A illustreert de opeenvolging van potentiële veranderingen die in de spiercel worden geregistreerd als gevolg van het stimuleren van het motoraxon., De pijl geeft het tijdstip aan waarop de schok naar de motor axon wordt gebracht. Merk op dat er een rustperiode is na de schok. De vertraging is te wijten aan de tijd die nodig is voor de actiepotentiaal in het motoraxon om zich te verspreiden vanaf de plaats van initiatie. Na de vertraging, zijn er twee soorten potentials geregistreerd in de spiercel. Ten eerste is er een relatief langzaam veranderend potentieel dat de focus zal zijn van de volgende discussie., Als die langzame initiële potentie voldoende groot is, zoals normaal in skeletspiercellen, wordt een tweede potentie, een actiepotentiaal, opgewekt in de spiercel.
figuur 4.3
actiepotentialen in skeletspiercellen zijn te wijten aan Ionische mechanismen die vergelijkbaar zijn met de eerder besproken mechanismen. In het bijzonder is er een spanningsafhankelijke verandering in Na+ permeabiliteit gevolgd door een vertraagde toename in K+ permeabiliteit. (Voor gladde spiercellen en hartspiercellen zijn de Ionische mechanismen echter verschillend.,)
de onderliggende gebeurtenis die de actiepotentiaal activeert kan worden onthuld door gebruik te maken van curare, een pijlgif dat door sommige Zuid-Amerikaanse Indianen wordt gebruikt. Een lage dosis curare (Deel B) vermindert de onderliggende gebeurtenis, maar is nog steeds niet voldoende verlaagd om onder de drempel te dalen. Als een iets hogere dosis curare wordt toegediend (deel C), wordt de langzame onderliggende gebeurtenis ondergrens. Het onderliggende signaal staat bekend als de endplate potential (EPP) omdat het een potentiële verandering is die aan de motor endplate wordt geregistreerd. Over het algemeen is het bekend als een prikkelend postsynaptisch potentieel (EPSP).,
Curare blokkeert het eindplaatpotentieel omdat het een competitieve remmer is van acetylcholine (ACh), de zender die vrijkomt in de presynaptische terminal. Curare blokkeert niet de spanningsafhankelijke na + – geleidbaarheid of de spanningsafhankelijke K+-geleidbaarheid die ten grondslag ligt aan het spieractiepotentiaal. Curare beïnvloedt de stimulus (de EPSP), wat normaal gesproken leidt tot het initiëren van het spieractiepotentiaal. Een dier dat is vergiftigd met curare zal verstikken omdat het proces van neuromusculaire overdracht aan de ademhalingsspieren is geblokkeerd.,
normaal gesproken is de grootte van de eindplaatpotentiaal vrij groot. De amplitude van de eindplaatpotentiaal is inderdaad ongeveer 50 mV, maar slechts ongeveer 30 mV is nodig om de drempelwaarde te bereiken. De extra 20 mV wordt de veiligheidsfactor genoemd. Daarom, zelfs als het eindplaatpotentiaal door vermoeidheid iets kleiner zou worden (bijvoorbeeld 40 mV in amplitude), zou de EPP de drempel bereiken en zou de één-op-één relatie tussen een actiepotentiaal in het motoraxon en een actiepotentiaal in de spiercel behouden blijven.
figuur 4.4
4.,3 verspreiding van de EPP
Wat zijn de eigenschappen van de EPP en hoe verhoudt deze zich tot de eigenschappen van het actiepotentieel?
is de eindplaatpotentiaal te wijten aan een spanningsafhankelijke verandering in na+ en K+ permeabiliteiten zoals de actiepotentiaal?
wordt het eindplaatpotentiaal gepropageerd op een alles-of-niets manier zoals het actiepotentiaal?
de figuur links illustreert een experiment dat de voortplanting van de eindplaatpotentiaal onderzoekt. De spiervezel wordt herhaaldelijk gespietst met elektroden met intervallen van 1 mm., (Merk op dat het eindplaatpotentiaal klein is omdat dit experiment wordt uitgevoerd in aanwezigheid van een lage concentratie curare zodat het eindplaatpotentiaal kan worden geregistreerd zonder de complicaties van het teweegbrengen van een actiepotentiaal.) Het endplate potentieel wordt niet gepropageerd op een alles-of-niets manier. Het verspreidt zich wel langs de spier, maar het doet het met decrement., Dus, de verspreiding van de endplate potential van zijn plaats van initiatie naar andere plaatsen langs de spiercel gebeurt passief en met vermindering, net zoals een subthreshold potentiële verandering in een deel van het axon zich langs het axon verspreidt, of net zoals een verandering in temperatuur op een punt op een metalen staaf zich langs de staaf verspreidt.
4.4 overzicht van de opeenvolging van gebeurtenissen die ten grondslag liggen aan de EPP
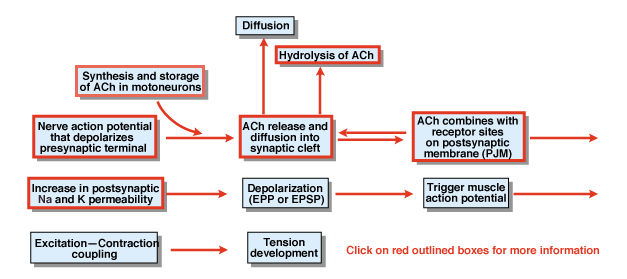
figuur 4.5
Wat zijn de andere stappen in het proces van chemische synaptische transmissie? Figuur 4.,5 geeft een overzicht. Een zenuwactiepotentiaal dat in het cellichaam van een ruggenmergmotorneuron wordt geïnitieerd verspreidt zich uit de ventrale wortels en dringt uiteindelijk de synaptische terminals van de motorneuronen binnen. Als gevolg van de actiepotentiaal, de chemische zender acetylcholine (ACh) wordt vrijgegeven in de synaptische spleet. ACh verspreidt zich over de synaptische spleet en bindt zich aan speciale receptoren op het postsynaptische of het postjunctionele membraan. De band van ACh aan zijn receptoren veroorzaakt een conformational verandering in een membraankanaal dat specifiek permeabel aan zowel Na+ als K+is., Als gevolg van een toename van na+ en K+ permeabiliteit, is er een depolarisatie van het postsynaptisch membraan. Die depolarisatie wordt de endplate potential of meer in het algemeen de EPSP genoemd. Als de EPSP voldoende groot is, zoals gewoonlijk bij de neuromusculaire junctie, leidt dit tot het initiëren van een actiepotentiaal in de spiercel. Het actiepotentiaal initieert het proces van excitatie contractie koppeling en de ontwikkeling van spanning. De duur van de endplate potential is ongeveer 10 msec.
twee factoren regelen de duur van de EPSP op de neuromusculaire junctie., Eerst wordt ACh verwijderd door diffusie. Ten tweede, een stof in de synaptische spleet, genaamd acetylcholinesterase (AChE), hydrolyseert of breekt ACh. AChE is een van de meest efficiënte enzymen die bekend zijn. Een enkel molecuul van AChE kan 600.000 molecules van ACh per minuut hydrolyseren.
4.5 rol van AChE
figuur 4.6
een belangrijke familie van stoffen, waaronder neostigmine, remt de werking van AChE. Neostigmine blokkeert de werking van AChE en maakt daardoor de eindplaatpotentiaal groter en langer van duur., Deze figuur illustreert twee endplate potentialen. Eén ervan werd geregistreerd in zoutoplossing en curare en een tweede na toevoeging van neostigmine aan de oplossing. (Curare wordt toegevoegd zodat de eigenschappen van de EPP kunnen worden bestudeerd zonder een actiepotentiaal in de spiercel te activeren.) Na het aanbrengen van neostigmine is het eindplaatpotentieel veel groter en langer van duur.
4.6 a Myasthenia Gravis
Myasthenia gravis wordt geassocieerd met ernstige spierzwakte vanwege een afname van het aantal acetylcholinereceptoren in de spiercel., Als de eindplaatpotentiaal kleiner is, zal de eindplaatpotentiaal de drempelwaarde niet bereiken. Als het niet lukt om de drempel te bereiken, zal er geen actie potentieel in de spiercel en geen samentrekking van de spier, die spierzwakte veroorzaakt. Neostigmine en andere remmers van AChE worden gebruikt voor de behandeling van patiënten met myasthenia gravis. Deze agenten maken de hoeveelheid acetylcholine die effectiever wordt vrijgegeven bereiken de resterende acetylcholine receptoren.
4.,6b Zenuwmiddelen
hoewel Ache-remmers een belangrijke therapeutische waarde hebben, zijn en worden sommige remmers nog steeds gebruikt als gifstoffen. Sommige Ache inhibitors zoals Soman en Sarin vormen een vrij onomkeerbaar blok van AChE. Dit blok leidt tot extreme niveaus van ACh in de synaptische spleet. Individuen zo vergiftigd sterven aan epileptische aanvallen en spierspasticiteit met inbegrip van ademhalingsspieren.
figuur 4.7
4.,Iontoforese van ACh
iontoforese is een interessante techniek die kan worden gebruikt om de hypothese dat ACh de neurotransmitterstof is op de neuromusculaire junctie verder te testen. Als ACh de zender is die door deze synaps wordt vrijgegeven, zou men voorspellen dat het mogelijk zou moeten zijn om kunstmatige toepassing van de zender te vervangen door de normale afgifte van de zender. Aangezien ACh een positief geladen molecuul is, kan het uit een micro-elektrode worden gedwongen om de versie van ACh van een presynaptic terminal te simuleren.
Figuur 4.,8
inderdaad, minieme hoeveelheden ACh kunnen worden toegepast op de omgeving van de neuromusculaire junctie. Figuur 4.8 vergelijkt een EPP geproduceerd door stimulatie van het motoraxon en de reactie op uitwerpingen van ACh. De potentiële verandering lijkt bijna identiek aan het eindplaatpotentieel dat door de normale versie van ACh wordt geproduceerd. Dit experiment biedt experimentele ondersteuning voor het concept dat ACh de natuurlijke zender is bij deze synaps.,
de respons op het uitwerpen van ACh heeft enkele andere interessante eigenschappen die allemaal consistent zijn met de cholinerge aard van de synaps bij de neuromusculaire kruising van het skelet. Neostigmine maakt de reactie op de iontoforese van ACh langer en groter. Curare vermindert de respons omdat het concurreert met de normale binding van ACh. Als ACh in de spiercel wordt uitgeworpen, gebeurt er niets omdat de receptoren voor acetylcholine niet in de binnenkant zitten; ze bevinden zich aan de buitenkant van de spiercel., De toepassing van acetylcholine op gebieden van de spier vanaf de eindplaat veroorzaakt geen reactie omdat de receptoren voor ACh bij het synaptische gebied worden geconcentreerd. om uw begrip tot nu toe te testen, overweeg hoe een agent zoals TTX de generatie van zowel een EPP als de reactie van een spiervezel op de iontoforetische toepassing van ACh zou beïnvloeden? TTX heeft geen effect op de reactie op ACh, maar het blokkeert wel de EPP. De reden waarom de reactie op ACh onaangetast is, is duidelijk, maar velen verwachten dat als er hier geen effect is, er ook geen effect op de EVP zou moeten zijn., Tetrodotoxine heeft geen invloed op de binding van acetylcholine aan de receptoren en heeft daarom geen invloed op de respons op directe toediening van ACh. Tetrodotoxine beïnvloedt echter het vermogen van een actiepotentiaal om in het motoraxon te worden opgewekt. Als een actiepotentiaal niet kan worden opgewekt in het motoraxon, kan dit niet leiden tot het vrijkomen van de zender. Zo zou tetrodotoxine de EPP volledig afschaffen. Het blok zou niet te wijten zijn aan een blok ACh receptoren, maar eerder aan een blok van één of andere stap voorafgaand aan de release van de zender.
4.,8 Ionische mechanismen van de EPP
Bernard Katz en zijn collega ‘ s waren pioniers in het onderzoeken van mechanismen van synaptische transmissie op de neuromusculaire junctie. Ze suggereerden dat het kanaal geopend door ACh was een die gelijke permeabiliteit voor zowel Na+ en K+had. Omdat het net zo doorlaatbaar was voor Na+ en K+, stelde Katz voor dat, als gevolg van de opening van deze kanalen, de membraanpotentiaal naar 0 mV zou gaan. (Een waarde van alfa in de GHK vergelijking gelijk aan één, die, wanneer gesubstitueerd in de vergelijking, een potentiaal van ongeveer 0 mV oplevert.,)
figuur 4.9
Het experiment in de figuur aan de linkerkant test dat concept. De spiercel is gepenetreerd met een opname elektrode evenals een andere elektrode die met een geschikte bron van potentieel kan worden verbonden om het membraanpotentieel kunstmatig te veranderen. Normaal gesproken is de membraanpotentiaal weer ongeveer -80 mV, een kleine hoeveelheid curare wordt toegevoegd zodat de EPP klein is. Katz merkte in deze experimenten dat de grootte van de EPP drastisch veranderde, afhankelijk van het potentieel van de spiercel., Als de membraanpotentiaal wordt verplaatst naar 0 mV, wordt er geen potentiële verandering geregistreerd. Als de membraanpotentiaal +30 mV wordt gemaakt, wordt de EPP omgekeerd. Dus drie verschillende stimuli produceren endplate potentialen die heel verschillend zijn van elkaar.
het ontbreken van een respons wanneer de potentiaal 0 mV bedraagt, is bijzonder informatief. Bedenk waarom er geen mogelijke verandering wordt geregistreerd. Vermoedelijk wordt de zender vrijgegeven en gebonden aan de receptoren., De eenvoudige verklaring voor een gebrek aan potentiële verandering is dat het potentieel waarop de opening van ACh-kanalen proberen te bereiken al is bereikt. Als de membraanpotentiaal positiever wordt gemaakt dan 0 mV, dan wordt de EPP omgekeerd. Het maakt niet uit wat de potentie is, de verandering in permeabiliteit heeft de neiging om het membraanpotentiaal naar 0 mV te verplaatsen! Als de rustpotentiaal negatiever is dan 0 mV, is er een opwaartse vervorming. Als het positiever is, is er een neerwaartse afbuiging. Als het al op 0 mV staat, is er geen doorbuiging.
Figuur 4.,10
dit potentieel wordt ook het omkeerpotentieel genoemd, omdat het het potentieel is waarbij het teken van het synaptische potentieel omkeert. Het experiment wijst erop dat, als resultaat van ACh band aan receptoren, specifieke kanalen even permeabel aan Na+ en K+worden. Deze verandering van permeabiliteit heeft de neiging om het membraanpotentieel van waar het zich aanvankelijk bevindt naar een nieuw potentieel van 0 mV te verplaatsen.
waarom bereikt het normale eindplaatpotentieel eigenlijk nooit 0 mV?, Een van de redenen is dat de opeenvolging van veranderingen in de permeabiliteit die ten grondslag liggen aan het actiepotentieel de veranderingen die door de EVP worden veroorzaakt, “wegvagen”. Maar zelfs als er geen actiepotentiaal zou worden geactiveerd, zou de EVP nog steeds geen 0 mV bereiken. Dit komt omdat de ACh kanalen slechts een kleine fractie van het totale aantal kanalen in spiervezels. De k + kanalen die een spiercel met zijn rustpotentieel begiftigen zijn ook aanwezig. Hun taak is om te proberen om de cel op het rustpotentieel te houden.,
het kanaal dat door ACh wordt geopend behoort tot een algemene klasse van kanalen die ligand-gated kanalen of ionotrope receptoren worden genoemd. Zoals in Figuur 4.10 wordt geïllustreerd, maakt de bindingsplaats van de zender deel uit van het kanaal zelf. Als resultaat van zender die aan de receptor binden (over het algemeen zijn twee molecules noodzakelijk), is er een conformational verandering in de proteã ne die een poriegebied toestaan te openen en ionen om onderaan hun elektrochemische gradiënten te stromen. Aanvullende details van het kanaal worden gepresenteerd in hoofdstuk 11.,
Test uw kennis
- opdracht 1
- A
- B
- C
- D
een eindplaatpotentiaal in een skeletspiercel kan in principe worden geproduceerd door een verminderde permeabiliteit voor welke van de volgende ionen? (Ga ervan uit dat er een eindige initiële permeabiliteit is voor elk van de hieronder genoemde ionen en dat er fysiologische concentratiegradiënten aanwezig zijn.):
A. Na+
B. Na+ en Ca2+
C. Ca2+
D., K +
een eindplaatpotentiaal in een skeletspiercel kan in principe worden geproduceerd door een verminderde permeabiliteit voor welke van de volgende ionen? (Ga ervan uit dat er een eindige initiële permeabiliteit is voor elk van de hieronder genoemde ionen en dat er fysiologische concentratiegradiënten aanwezig zijn.):
A. Na+ dit antwoord is onjuist.
Een eindplaatpotentiaal is een depolarisatie die normaal wordt veroorzaakt door de gelijktijdige toename van de permeabiliteit voor natrium-en kaliumionen., Als er een selectieve afname van de natriumpermeabiliteit was, zou een dergelijke afname niet leiden tot een depolarisatie. Eerder, zou het eigenlijk leiden tot een hyperpolarisatie omdat alpha in de GHK vergelijking zou worden verminderd. De verhouding van de permeabiliteit tot natrium en kalium zou worden begunstigd in de richting van de kaliumpermeabiliteit, waardoor de membraanpotentiaal naar de potentiaal van het kaliumevenwicht wordt verplaatst en een hyperpolarisatie wordt veroorzaakt.
B. Na+ en Ca2 +
C. Ca2 +
D., K +
een eindplaatpotentiaal in een skeletspiercel kan in principe worden geproduceerd door een verminderde permeabiliteit voor welke van de volgende ionen? (Ga ervan uit dat er een eindige initiële permeabiliteit is voor elk van de hieronder genoemde ionen en dat er fysiologische concentratiegradiënten aanwezig zijn.):
A. Na+
B. Na+ en Ca2+ dit antwoord is onjuist.
indien de permeabiliteit voor natrium en calcium zou afnemen, zouden de gevolgen ervan vergelijkbaar zijn met die van keuze A. Indien alleen de natriumpermeabiliteit zou worden verminderd, zou de cel geneigd zijn te hyperpolariseren., Op dezelfde manier kan het verminderen van de permeabiliteit voor calcium de cel ook hyperpolariseren. Het calciumevenwicht potentieel is een zeer positieve waarde, en als er enige tonische rustpermeabiliteit voor calcium zou zijn, zou die permeabiliteit bijdragen tot een tonische depolarisatie van het membraanpotentieel. Vandaar, zou een afname van de calciumdoorlaatbaarheid dat tonische depolariserende effect verwijderen en resulteren in een hyperpolarisatie.
C. Ca2 +
D., K +
een eindplaatpotentiaal in een skeletspiercel kan in principe worden geproduceerd door een verminderde permeabiliteit voor welke van de volgende ionen? (Ga ervan uit dat er een eindige initiële permeabiliteit is voor elk van de hieronder genoemde ionen en dat er fysiologische concentratiegradiënten aanwezig zijn.):
A. Na +
B. Na+ en Ca2 +
C. Ca2+ dit antwoord is onjuist.
Een afname van de calciumdoorlaatbaarheid alleen zou, als er al iets was, een hyperpolarisatie veroorzaken. Zie logica van de respons op keuze B.
D., K +
een eindplaatpotentiaal in een skeletspiercel kan in principe worden geproduceerd door een verminderde permeabiliteit voor welke van de volgende ionen? (Ga ervan uit dat er een eindige initiële permeabiliteit is voor elk van de hieronder genoemde ionen en dat er fysiologische concentratiegradiënten aanwezig zijn.):
A. Na+
B. Na+ en Ca2+
C. Ca2+
D. K+ dit antwoord is CORRECT!
Een afname van de kaliumpermeabiliteit zou leiden tot een depolarisatie vergelijkbaar met een eindplaatpotentiaal., Dit is zo omdat er in rust een tonische permeabiliteit voor kalium en natrium. De hoge doorlaatbaarheid voor kalium heeft de neiging om het membraanpotentiaal dicht bij het potentiaal van het kaliumevenwicht te houden. Als die rustpermeabiliteit wordt verminderd, zou alfa in de Goldman-vergelijking een grotere waarde worden, waardoor het membraanpotentiaal een beetje dichter bij het natriumevenwicht potentiaal wordt verplaatst (d.w.z. een depolarisatie).