Inleiding
Darwin ‘ s grote inzicht, en het verenigende principe van de biologie vandaag, is dat alle soorten verwant zijn aan elkaar zoals zussen, neven en verre verwanten in een enorme stamboom van het leven. De implicaties zijn adembenemend; als we ver genoeg terug in de tijd konden reizen, zouden we gemeenschappelijke voorouders vinden tussen onszelf en elk ander levend organisme, van stekelvarkens tot flamingo ‘ s tot Cactussen., Onze directe evolutionaire familie bestaat uit de mensachtigen, de groep primaten die de “kleine apen” (siamangs en gibbons) en de “grote apen” (chimpansees, bonobo ‘s, gorilla’ s en orang-oetans) omvat. Bij de mensapen zijn onze naaste verwanten de chimpansees en bonobo ‘ s (Figuur 1). Het fossielenbestand, samen met studies van menselijk en aap DNA, geven aan dat de mens een gemeenschappelijke voorouder deelde met chimpansees en bonobo ‘ s ergens ongeveer 6 miljoen jaar geleden (mya)., We beginnen deze discussie over de evolutie van onze soort in Afrika, tegen het einde van de geologische periode bekend als het Mioceen, net voordat onze afstamming afwijkt van die van chimpansees en bonobo’ s.
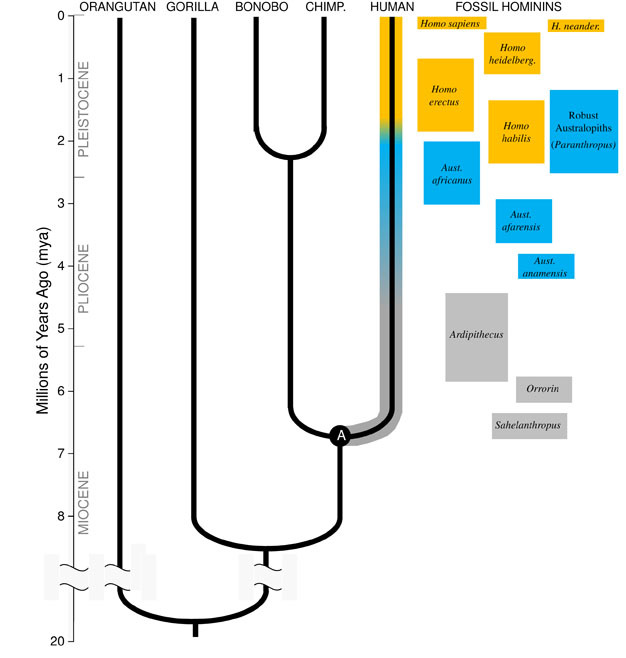
Mioceen oorsprong van de Hominine lijn
om de evolutie van een soort te begrijpen, moeten we eerst de voorouderlijke toestand ervan vaststellen: van welk soort dier is het geëvolueerd? Voor onze afstamming vereist dit dat we proberen de laatste gemeenschappelijke voorouder van mensen en chimpansees te reconstrueren (gemarkeerd met “A” in Figuur 1). De mens-chimpansee laatste gemeenschappelijke voorouder (HC-LCA)is de soort waarvan de hominine lijn en de chimpansee & bonobo lijn diverged., Homininen zijn soorten op onze tak van de hominoïde boom na de splitsing met de chimpansee & bonobo lijn, inclusief alle uitgestorven soorten en evolutionaire zijtakken (figuur 1).er was een grote diversiteit aan apensoorten in het Mioceen, met tientallen soorten bekend uit het fossielenbestand in Afrika, Europa en Azië. Deze soorten varieerden in hun anatomie en ecologie, en het is niet duidelijk welke, indien aanwezig, van de fossiele soorten tot nu toe de HC-LCA (Kunimatsu et al. 2007; Young and MacLatchy, 2004)., Toch weten we uit fossiel en vergelijkend bewijs dat het veel meer leek op levende apen dan op levende mensen. De HC-LCA zou een brein en lichaam ter grootte van een aap hebben gehad, met relatief lange armen en vingers en een grijpende voet die het mogelijk maakte om in de bomen te foerageren. De hoektanden waren waarschijnlijk groot en scherp, zoals te zien is bij verschillende Mioceen hominoïden. Bovendien waren de hoektanden waarschijnlijk seksueel dimorf, met mannetjes met veel grotere hoektanden dan vrouwtjes, zoals gezien onder de levende mensapen en Mioceen fossielen., Net als levende apen zou hij viervoetig (op handen en voeten) op de grond gelopen hebben, en zijn dieet zou bijna volledig uit plantaardig voedsel bestaan, voornamelijk fruit en bladeren.
vroege Homininen
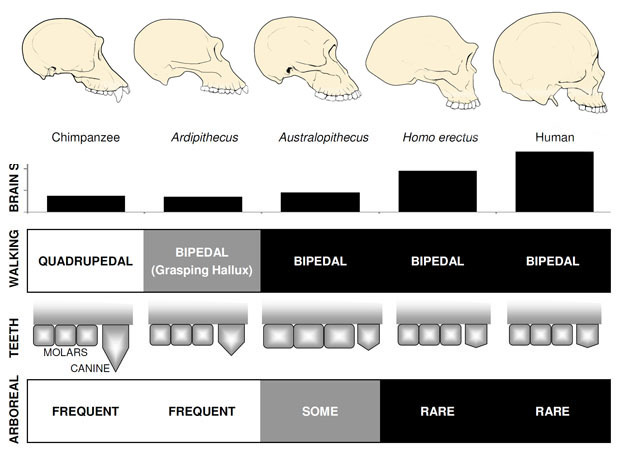
veranderingen van een aapachtige anatomie zijn waarneembaar in hominoïde fossielen uit het late Mioceen in Afrika. Sommige hominoïde soorten uit deze periode vertonen kenmerken die typerend zijn voor de mens, maar niet worden gezien in de andere levende apen, waardoor paleoantropologen concluderen dat deze fossielen vroege leden van de hominine lijn vertegenwoordigen., De eerste menselijke kenmerken die in het fossielenbestand van de mens voorkomen, zijn tweevoetige wandelingen en kleinere, stompe hoektanden. de oudste bekende homininen zijn Sahelanthropus tchadensis uit Tsjaad (Brunet et al. 2005) en Orrorin tugenensis uit Kenia (Senut et al. 2001). Sahelanthropus, gedateerd tussen 6 en 7 mya, is bekend van een grotendeels complete schedel en enkele andere fragmentarische overblijfselen. De grootte van de hersenen, 360cc, ligt binnen het bereik van chimpansees, en de schedel heeft een enorme wenkbrauwkam, vergelijkbaar in dikte met mannelijke gorilla ‘ s (Brunet et al. 2005)., Echter, de positie en oriëntatie van het foramen magnum, het gat in de basis van de schedel waardoor het ruggenmerg passeert, suggereert dat Sahelanthropus stond en liep tweevoetig, met zijn wervelkolom verticaal gehouden zoals in de moderne mens in plaats van horizontaal zoals in apen en andere viervoeters (Zollikofer et al. 2005). Orrorin is vooral bekend van postcraniale fossielen, waaronder een partieel dijbeen. Het proximale deel van het dijbeen vertoont overeenkomsten met die van de moderne mens, wat suggereert dat de soort tweevoetig was (Pickford et al. 2002)., Er zijn geen schedels van Orrorin gevonden, dus de schedelmorfologie en hersengrootte zijn onzeker. In zowel Orrorin als Sahelanthropus zijn de hoektanden van mannetjes groter en puntiger dan bij de moderne mens, maar zijn ze klein en stomp in vergelijking met de hoektanden van mannelijke apen. Dit suggereert dat het seksuele dimorfisme van honden — en bij uitbreiding, de concurrentie tussen mannetjes om toegang te krijgen tot de vrouwtjes — bij deze vroege mensapen verminderd was in vergelijking met de mensapen.de bekendste vroege hominine is Ardipithecus ramidus, een 4.,4 miljoen jaar oude soorten uit Ethiopië, die bekend is van een bijna compleet skelet en tal van andere tand-en skeletresten (White et al. 2009). Ar. ramidus en een oudere, verwante soort bekend van fragmentarische resten, Ar. kadabba (5.8-5.2 mya), hebben gereduceerde hoektanden vergelijkbaar met die van Orrorin en Sahelanthropus. De schedel van Ar. ramidus is vrij aapachtig en in grote lijnen vergelijkbaar met die van Sahelanthropus, met een klein chimpansee-formaat hersenen van 300–350cc (Figuur 2). Het Ardipithecus postcraniaal skelet is intrigerend., Hoewel het bekken sterk gefragmenteerd is, vertoont het een morfologie die heel anders is dan die van levende apen, met een kortere, meer komvormige vorm die sterk suggereert dat Ardipithecus tweevoetig liep; dit komt overeen met de positie van het foramen magnum, wat wijst op een rechtopstaande houding. Echter, zijn lange voorpoten en vingers en zijn divergerende, grijpende eerste teen (hallux) suggereren Ardipithecus bracht veel van zijn tijd in de bomen. De algemene indruk is van een grotendeels boombewonende soort die tweevoetig liep wanneer hij zich op de grond waagde.,
Australopithecus
rond 4mya vinden we de vroegste leden van het geslacht Australopithecus, homininen die bedreven terrestrische tweevoeters waren, maar de bomen bleven gebruiken voor voedsel en bescherming. De eerste exemplaren van Australopithecus werden ontdekt in Zuid-Afrika in 1924 (dart, 1925), en onderzoek in de daaropvolgende acht decennia hebben honderden fossielen geproduceerd van verschillende soorten op locaties in Oost-en Zuidelijk Afrika., We weten nu dat Australopithecus een zeer succesvol geslacht was dat bijna drie miljoen jaar standhield (figuur 1).de bekendste Australopithecus-soorten zijn A. afarensis (3,6–2,9 mya) uit Oost–Afrika en A. africanus (3,2-2,0 mya) uit Zuid-Afrika., Het bekken en de onderste ledematen van deze soorten geven duidelijk aan dat ze volledig tweevoetig waren: het bekken is kort en komvormig, waardoor de bilspieren naar de zijkant van het lichaam worden gebracht, zoals bij de moderne mens, voor stabilisatie van de romp tijdens tweevoetig, en de eerste teen is in lijn met de andere tenen (Ward, 2002; Harcourt-Smith and Aiello, 2004). De voet van de Australopithecus kan zelfs een menselijke boog hebben gehad, gebaseerd op analyse van de middenvoetsbeentjes en de gefossiliseerde voetafdrukken van Laetoli (Ward et al. 2011)., In vergelijking met de moderne mens waren de onderarmen echter lang en de vingers en tenen lang en enigszins gebogen, wat erop wijst dat Australopithecus de bomen regelmatig gebruikte om te foerageren en misschien ‘ s nachts als toevluchtsoord voor roofdieren. Deze gemengde terrestrische& arboreale strategie zou deze soorten goed hebben gediend in het gemengde bos-en savannemilieu waarin ze leefden.
hersengrootte in Australopithecus varieerde tussen 390 en 515cc, vergelijkbaar met chimpansees en gorilla ‘ s (Falk et al., 2000), wat erop wijst dat cognitieve vaardigheden in grote lijnen vergelijkbaar waren met levende apen (Figuur 2). De lichaamsgrootte in Australopithecus was vrij klein en seksueel dimorf, ongeveer 30 kg voor vrouwen en 40 kg voor mannen (McHenry, 1992). Dit niveau van dimorfisme wordt niet weerspiegeld in de hoektanden, die klein, stomp en monomorf waren zoals in eerdere homininen. in tegenstelling tot de hoektanden waren de molaire tanden in Australopithecus veel groter dan die van eerdere homininen en hadden ze dikker email. Dit suggereert hun dieet opgenomen harde, lage kwaliteit plantaardige voedingsmiddelen die krachtige kauwen nodig om te verwerken., Een subgroep van Australopithecus, bekend als de” robuuste ” australopieten (vaak aangeduid door een apart geslacht Paranthropus) vanwege hun enorme tanden en kauwspieren, nam deze aanpassing tot het uiterste. De meeste Australopithecus soorten waren uitgestorven door 2 mya, maar sommige robuuste vormen bleven bestaan tot ongeveer 1,2 mya in Oost-en Zuid-Afrika.het geslacht Homo
De vroegste fossielen van ons eigen geslacht, Homo, worden gevonden in Oost-Afrika en gedateerd op 2,3 mya (Kimbel et al. 1997)., Deze vroege exemplaren zijn in hersenen en lichaamsgrootte gelijkaardig aan Australopithecus, maar tonen verschillen in hun molaire tanden, die een verandering in dieet suggereren. Inderdaad, met ten minste 1,8 miljoen jaar gebruikten vroege leden van ons geslacht primitieve stenen werktuigen om karkassen van dieren te slachten, en voegden energierijk vlees en beenmerg toe aan hun plantaardige voeding.
het oudste lid van het geslacht Homo, H. habilis (2.3-1.4 mya) wordt gevonden in Oost-Afrika en wordt geassocieerd met afgeslachte dierlijke botten en eenvoudige stenen werktuigen (Blumenschine et al. 2003). Zijn meer formidabele en wijdverspreide afstammeling, H., erectus, wordt gevonden in heel Afrika en Eurazië en bleef bestaan van 1,9 mya tot 100 kya, en misschien zelfs later (Anton, 2003). Net als de moderne mens miste H. erectus de aanpassingen van de voorpoot om te klimmen die in Australopithecus te zien zijn (figuur 2). Zijn globale expansie suggereert H. erectus ecologisch flexibel was, met de cognitieve capaciteit om zich aan te passen en te gedijen in enorm verschillende omgevingen. Niet verrassend, het is met H. erectus dat we een grote toename van de hersengrootte beginnen te zien, tot 1.250 cc voor latere Aziatische specimens (Anton, 2003). Molaire grootte wordt verminderd in H., erectus ten opzichte van Australopithecus, als gevolg van zijn zachtere, rijkere dieet.
rond 700 kya, en misschien eerder, gaf H. erectus in Afrika aanleiding tot H. heidelbergensis, een soort die erg op ons leek in termen van lichaamsverhoudingen, tandheelkundige aanpassingen en cognitieve vaardigheden (Rightmire, 2009). H. heidelbergensis, vaak aangeduid als een” archaïsche ” Homo sapiens, was een actieve jager op groot wild, produceerde geavanceerde Levallois-stijl gereedschappen, en door ten minste 400 kya had geleerd om vuur te beheersen (Roebroeks and Villa, 2011). Neanderthalers (H., neanderthalensis), cold-adapted hominins met stevige lichaamsbouw, complex gedrag, en hersenen vergelijkbaar in grootte aan de Onze, worden verondersteld te zijn geëvolueerd uit H. heidelbergensis populaties in Europa door ten minste 250 kya (Rightmire, 2008; Hublin, 2009).fossiel en DNA-bewijs suggereert dat onze eigen soort, H. sapiens, geëvolueerd is in Afrika 200 kya (Relethford, 2008; Rightmire, 2009), waarschijnlijk van H. heidelbergensis. De verhoogde gedragsmatige verfijning van H., sapiens, zoals aangegeven door onze grote hersenen (1400 cc) en archeologische bewijzen van een bredere tool set en slimme jachttechnieken, liet onze soort om te bloeien en te groeien op het Afrikaanse continent. Door 100kya, onze soort gemorst in Eurazië, uiteindelijk uit te breiden over de hele wereld in Australië en de Amerika ‘ s (DiGiorgio et al. 2009). Onderweg verplaatste onze soort andere mensachtigen die ze tegenkwamen, waaronder Neanderthalers in Europa en soortgelijke vormen in Azië. (Merk op dat niet iedereen het eens is met deze interpretatie van de gegevens, Zie Tryon en Bailey)., Studies van oud DNA geëxtraheerd uit Neanderthaler fossielen suggereren dat onze soort af en toe met hen heeft gekruist (Green et al., 2010). Onze toenemende wereldwijde impact zet zich vandaag voort, aangezien culturele innovaties zoals landbouw en verstedelijking het landschap en de soorten om ons heen vormen.
samenvatting
de evolutie van onze soort uit een aapachtige Mioceen voorouder was een complex proces. Onze afstamming is vol met zijtakken en evolutionaire doodlopende sporen, met soorten zoals de robuuste australopieten die meer dan een miljoen jaar stand hielden voordat ze vervaagden., Sommige menselijke eigenschappen, zoals bipedalisme, evolueerden heel vroeg, terwijl andere, zoals grote hersenen, pas relatief recent evolueerden. Nog andere eigenschappen, zoals de molaire grootte, evolueerden in één richting en werden later teruggeduwd door ecologische druk te veranderen. In plaats van een krachtig schip dat een rechte koers in de richting van een vooraf bepaalde bestemming zet, past de evolutie van onze afstamming-inderdaad, van de afstamming van welke soort dan ook — bij het beeld van een reddingsboot die door de veranderende zeeën van milieuveranderingen, genetisch geluk en geologische toeval heen en weer geslingerd wordt. Je vraagt je af waar de komende zes miljoen jaar ons heen kunnen brengen.