Inleiding
een manier om functionele corticale organisatie te beschrijven is door middel van kaarten die de gehele cortex in kleine gebieden verpakken, elk met specifieke hersenfuncties. De meest populaire kaart werd voor het eerst beschreven door Brodmann (1909) die de cortex van elk halfrond in 52 gebieden verdeelde., Deze gebieden zijn gesegmenteerd op basis van histologische verschillen, en hebben geen overeenstemming met de anatomische sub-divisies van de hersenen in kwabben en gyri. Brodmanns gebied 44 (BA44) is een van de weinige functionele gebieden die een precieze overeenkomst heeft met een van de anatomische onderverdelingen van de cortex. BA44 is inderdaad beperkt en beperkt door pars opercularis van de linker inferieure frontale gyrus, de kern van de expressieve taalfunctie beschreven door Broca (1861).,
Het is lastig om een dergelijke verscheidenheid aan functies uit te leggen vanuit een segregationistisch model van hersenfuncties gebaseerd op modules. De meeste moderne modellen pleiten voor multimodulaire benaderingen die cognitie verklaren als een variantie in netwerkconfiguratie. Dit betekent dat elk specifiek gebied (bijvoorbeeld BA44) kan verbinden met verschillende modules, afhankelijk van de taak, wat specifieke netwerkconfiguraties oplevert die verantwoordelijk zijn voor een bepaalde functie. Hersenconnectiviteit, de term om naar deze visie te verwijzen, kan betere complexe cognitieve, gedragsmatige en neuropsychologische verschijnselen verklaren dan eenvoudige lokalisatiemodellen., De neurale netwerkkarakterisatiemodellen zijn van waarde in de huidige status van cognitieve neurowetenschappen, en om deze queeste te bereiken zijn input van verschillende methodes noodzakelijk.
als gevolg hiervan is er nieuw gevonden interesse in hersenconnectiviteit door de komst van diffusion tensor imaging (DTI) en resting-state functional MRI (fMRI). DTI is in staat om vezelgebieden van levend neuronaal weefsel te identificeren met behulp van een recent ontwikkelde techniek genaamd tractografie., Tractografie is een imaging post-processing techniek die minuscule waterdiffusie trajecten (tensoren) fuseert in een keten die neurale traktaten vertegenwoordigt. Rust-staat fMRI is een andere vorm van het berekenen van post-processing procedure die hersenverbindingen afbeeldt door het vertegenwoordigen van discrete hersengebieden waarvan spontane oscillaties zijn synchroon. Deze twee nabewerkingsprocedures zijn momenteel de populairste methoden om hersenconnectiviteit te bestuderen.
nieuwe terminologie is gecreëerd om de bevindingen of techniek te definiëren die geassocieerd zijn met moderne hersenconnectiviteitsstudies., (1) structurele connectiviteit, verwijst naar de afbeelding van vezels door tractografie; (2) functionele connectiviteit, verwijst naar kaarten van synchronische hersenen oscillaties, (3) effectieve connectiviteit, verwijst naar taak-gebaseerde fMRI waarin statistische en heuristische benaderingen beoordelen de richting van de gegevensstroom in de geactiveerde modules van fMRI., Tractografie kan deterministisch of probabilistisch zijn; functionele connectiviteit kan gebaseerd zijn op onafhankelijke Componentanalyse (Ica) van de volledige gegevens, of beperkter met een weergave van remote synchrony gerelateerd aan de gemiddelde signaalvariatie van een bepaald gebied van belang (ook bekend als op seeded gebaseerde functionele connectiviteit). Effectieve connectiviteit kan worden onderzocht met ten minste twee vrij complexe wiskundige benaderingen .
hoewel al deze methoden vergelijkbare resultaten kunnen opleveren, kunnen ze ook verschillen., Bijvoorbeeld, het gebruik van deterministische tractografie heeft beperkte resolutie met kruising vezels, terwijl probabilistische tractografie lijkt om de handel gevoeligheid voor specificiteit (Yo et al., 2009). Functionele connectiviteit wordt waargenomen tussen regio ‘ s waar er weinig of geen structurele verbinding is (Damoiseaux en Greicius, 2009). Echter, functionele connectiviteit kan veranderen en kan worden beïnvloed door snel leren, training van een taak specifieke prestaties of een laesie (Kelly en Garavan, 2005; Hasenkamp en Barsalou, 2012; Jolles et al., 2013; Vahdat et al.,, 2014), terwijl de structurele connectiviteit stabieler is en de veranderingen hoofdzakelijk met laesies verband houden. De overgrote meerderheid van de studies van functionele connectiviteit zijn gebaseerd op rust-state fMRI. Weinig studies hebben hersenconnectiviteit onderzocht tijdens of na taken (bijv., Caclin en Fonlupt, 2006; Bernal et al., 2013; Dima et al., 2013). De kaarten van de hersenenactivering in taak-gerelateerde fMRI hebben connectiviteit getoond maar altijd beperkt tot de onderzochte functie. Dit is het gevolg van effectieve connectiviteit.,
een recent beschreven methodologie gebaseerd op een vrij beperkte meta-analysetechniek is ook beschreven om functionele netwerken van de hersenen weer te geven. De methode is genoemd meta-analytic connectivity model (MACM; Bzdok et al., 2013; Ardila et al., 2014; Kohn et al., 2014). In de huidige studie gebruiken we deze methode om Broca ‘ s area core netwerk betrokken bij expressieve taal te beschrijven. We hebben de voorkeur gegeven aan de term “Pooling-data connectivity study” om verwarring met de standaard meta-analyse methodologie die meestal bredere bronnen vereist te voorkomen.,
materialen en methoden
de gegevensbron voor deze pooling-data connectiviteit studie was brainmap.org de reden om alleen deze bron te gebruiken is dat deze database specifieke en gesystematiseerde velden van informatie en software-specifieke codering van activeringscoördinaten biedt die nauwkeurige, automatische en consistente selectie van het te bestuderen Monster mogelijk maken. De output van de database wordt ook gelezen door een specifieke software, ook geleverd door brainmap.org. dus, de database van Brainmap1 werd benaderd met behulp van Sleuth 2.,2, open software die door dezelfde website, op 20 augustus, 2014 wordt verstrekt. De zoekomstandigheden waren: (1) studies waarin melding werd gemaakt van ba44 of Broca ‘ s activering van het gebied; (2) studies waarbij gebruik werd gemaakt van fMRI; (3) normale proefpersonen; (4) activeringen: “alleen activering” (verwijderingsrapport van de-activeringen); (5) rechtshandige proefpersonen; (6) leeftijd 20-60 jaar; (7) domein: taal. De zoekopdracht wordt automatisch uitgevoerd door Sleuth, het geven van een lijst van papers die voldoen aan de selecton criteria., Personen ouder dan 60 werden uitgesloten om twee redenen, (1) leeftijd wordt meestal beschouwd als een sterke risicofactor voor cognitieve achteruitgang in het algemeen, en dementie in het bijzonder (Ritchie en Kildea, 1995; Ferri et al., 2006); (2) enige verbale vermogensafname wordt waargenomen na de leeftijd van 60 jaar (bijv., Alwin and McCammon, 2001; Ardila, 2007); bepaalde verbale vaardigheden, zoals verbale vloeiendheid –een taalproductievermogen waarbij BA44 betrokken is – neemt duidelijk af na deze leeftijd (Tombaugh et al., 1999).
negenenzestig papers met 102 van de 407 experimenten kwamen overeen met de criteria. Op dit moment werden uitsluitingscriteria toegepast., Artikelen werden uitgesloten als ze tweetalige onderwerpen hadden of taken in Oosterse talen hadden uitgevoerd, taken beperkt tot automatische spraak, taken beperkt tot ontvankelijke taal. Papers die patiënten rapporteerden of papers waarin taalbetrokkenheid marginaal of bijkomstig was van andere cognitieve taken (geheugen, aandacht, remming) of gemedieerde reacties van de proefpersoon werden ook uitgesloten. Onderwerpen van beide geslachten werden opgenomen. Op basis van deze uitsluitingscriteria werden 12 artikelen uitgesloten., Zo werden de fMRI-resultaten van 57 papers samengevoegd voor verdere analyse met 883 van de 914 proefpersonen, 84 van de 338 experimenten, 175 van de 280 condities en 1247 van de 3699 locaties (Tabel 1). Activeringen geassocieerd met BA44 (zoekcriteria) werden automatisch verkregen uit de Sleuth software. Dit automatische rapport lijst een aantal clusters gedefinieerd door het centrum van massa (in MNI-coördinaten), clustervolume in mm3, en intensiteit. Deze coördinaten, per onderwerp / taak / papier werden geëxporteerd als tekstbestanden (gepoolde resutls) voor analyse op de volgende stap.,
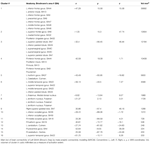
tabel 1. Activeringswaarschuwingsschatting (ALE) rapport.
de statistische significantie van clusters die op de gepoolde gegevens werden gevonden, werd vervolgens geanalyseerd met behulp van de activeringswaarschijnlijkheidsschatting (ALE)-methode. Deze stap werd uitgevoerd met de open source software GingerALE2. ALE is een methode om op coördinatie gebaseerde hersenactivaties te analyseren in pooling-data studies. De beschrijving van de wiskunde van ALE gaat het doel van dit rapport te boven., In een notendop, ALE behandelt gerapporteerde pieken van activering als ruimtelijke kansverdelingen gecentreerd op de gegeven coördinaten. ALE berekent de Vereniging van activeringswaarschijnlijkheden voor elke voxel, waardoor onderscheid kan worden gemaakt tussen echte convergentie van activeringshaarden en willekeurige clustering (ruis). ALE-scores verkregen uit duizenden willekeurige iteraties worden gebruikt om p-waarden toe te wijzen aan de waargenomen clusters van activering. Voor meer informatie over de theorie van ALE wordt de lezer geadviseerd om het werk van Eickhoff et al te lezen. (2009). Onze ALE-kaarten waren drempelwaarde bij p < 0.,01 gecorrigeerd voor meerdere vergelijkingen met false discovery rate. Alleen clusters van 200 of meer kubieke mm indien als geldige clusters aanvaard. ALE resultaten werden overlayed op een anatomische template geschikt voor MNI coördinaten, ook geleverd door brainmap.org voor dit doel hebben we gebruik gemaakt van de Multi-Image Analysis GUI (Mango)3. Een mozaïek van 3 × 3 transverale insets van fusion beelden werd verkregen met behulp van dezelfde tool, het selecteren van elke 3-4 beelden te beginnen op afbeelding nr. 10, en geëxporteerd naar een 2D-jpg-afbeelding.
resultaten
zestien significante clusters van activering werden gevonden met de ALE-procedure., Tabel 1 toont deze clusters gerangschikt op hun volume in kubieke millimeters. De belangrijkste cluster omvat BA44 en de aangrenzende gebieden. Dit zijn de voorste insula, de inferieure en middelste frontale gyri en de pre-centrale gyrus. De tweede Cluster wordt gevestigd in de linker pre-SMA en anterior cingulate gyrus die BA6 en 32 impliceren. De betrokkenheid van de rechter anterieure cingulate gyrus zou echt kunnen zijn of, hoogstwaarschijnlijk, een effect van het gladmaken van de naburige contralaterale homologe structuur., De derde cluster bevindt zich in de linker bovenste en onderste pariëtale lobule, een gebied dat gedeeld wordt door BAs 7, 39 en 40. De vierde cluster omvatte een aantal spiegelgebieden van de linker Broca (rechter BA44, rechter anterior insula en rechter BA9) en een subcorticale structuur, het putamen. De vijfde cluster betrof de linker spoelvormige gyrus. De zesde cluster vertegenwoordigt de kern van het ontvankelijke taalgebied of Wernicke ‘ s gebied. De volgende cluster van belang was gelegen in de linker thalamus., Negen andere clusters werden vermeld in het geautomatiseerde rapport van GingerAle, zoals opgesomd in Tabel 1, Zij zijn de linker putamen, de rechter pariëtale kwab, de occipitale kwab, het cerebellum, en de rechter precentrale gyrus.
discussie
We presenteren Broca ‘ s gebiedsnetwerk specifiek voor taaltaken met behulp van een methode van ALE in gepoolde fMRI-gegevens. Onze methode verschilt van anderen door de connectiviteit van een specifiek gebied in zijn breedste waaier van potentialiteit af te beelden, door zich te concentreren in de karakterisering van specifieke netwerken die een specifiek cognitief domein of functie subserveren.,
We ontdekten dat het taalnetwerk van BA44 uit 16 clusters bestaat. De eerste zes clusters van activering zijn gevestigde taalgebieden: cluster 1 vertegenwoordigt de linker infero-laterale frontale gyrus en de anterior insula, die samen in de literatuur zijn erkend als het expressieve taalgebied (Benson and Ardila, 1996)., De grote omvang van de cluster onthult de dichte naburige connectiviteit om gebieden grenzend aan BA44 door u vezels; cluster 2, vertegenwoordigt de aanvullende Motor gebied, waaraan de prior structureel is verbonden door de aslant frontale fasciculus beschreven door Catani et al. (2013), en waarschijnlijk geassocieerd met verbale vloeiendheid en initiatie van spraak (Martino et al., 2012); cluster 3, vertegenwoordigt de activering van de linker superieure en inferieure pariëtale lobule, direct of indirect verbonden met vezels van de boogvormige fasciculus of de inferieure occipitofrontale fasciculus (Dick et al.,, 2013). De verbinding met deze pariëtale gebieden zijn meer waarschijnlijk gerelateerd aan de toegang tot verbale werkgeheugen knooppunten (Jonides et al., 1998); cluster 4, toonde activering van homologe gebieden van cluster 1; cluster 5 en 6 waren gerelateerd aan canonieke Wernicke ‘ s Gebieden, hoogstwaarschijnlijk verbonden door de boogvormige fasciculus en subserving fonologische transferering functies (Dick et al., 2013).
Clusters 7 tot 16 bestonden uit activering van subcorticale gebieden, mediale dorsale kern van de linker thalamus; linker putamen en rechter cerebellum., De betrokkenheid van het linker cerebellum is twijfelachtig en zal later worden behandeld. Andere kleine clusters bevinden zich in contralaterale homologe gebieden van activeringen (BA7 en BA22), secundaire visuele gebieden (BA18 en BA19) in beide hemisferen en activering van de rechter precentrale gyrus. De betrokkenheid van deze gebieden wordt niet goed begrepen., Visuele gebieden kunnen worden betrokken bij verbale taken als het Onderwerp” re-visualiseer ” objecten en scènes beschreven door het verbale materiaal; precentrale gyrus kan worden betrokken als een consecuence van subvocalisatie, die aanwezig kan zijn wanneer blootgesteld aan verbale materiaal als een strategie voor repetitie beelden met motor aanwijzingen (Smith et al., 1995). Van belang is het gebrek aan connectiviteit met links BA45 en links BA21, niet vermeld in het ALE-automatic tekstrapport, noch weergegeven in de weergave afbeelding. Het gebrek aan betrokkenheid van links BA45 en BA39 zal later worden aangepakt.,
voor zover wij weten zijn er geen eerdere pogingen gemeld om de functionele connectiviteit van Broca ‘ s gebied in taal vast te stellen. Slechts weinig publicaties hebben echter studies gerapporteerd die de hersenconnectiviteit met betrekking tot specifieke taken of taalfuncties beoordelen. Met behulp van een seed-based rust-state functionele connectiviteitsanalyse Zhu et al. (2014) hebben aangetoond het taalnetwerk zaaien Broca ’s en Wernicke’ s Gebieden. Ze toonden aan dat Broca ‘ s lateraal achtergelaten werden., Ook, met behulp van zaad-gebaseerde rust-staat fMRI in een cohort van 970 gezonde proefpersonen Tomasi en Volkow (2012) gevonden activering van de canonieke prefrontale, temporale en pariëtale regio ‘ s, bilaterale caudate en links putamen/globus pallidus, en subthalamische kern. De auteurs gebruikten zowel Wernicke ’s als Broca’ s gebieden als zaaiende regio ‘ s. Er zijn ook enkele studies van structurele connectiviteit van Broca ‘ s gebied. Morgan et al. (2009) rapporteerde het combineren van DTI en rust-state functionele connectiviteit om de connectiviteit tussen SMA en expressieve taalgebieden te beoordelen., In een meta-analytische studie van Eickhoff et al. (2009) beschreef het expressieve netwerk door het bundelen van resultaten van vloeiende taken en het uitvoeren van DCM (Heim et al., 2009). Ze vonden het kernnetwerk bestaande uit BA44, anterior insula, BA6 (premotor cortex), en BA4 (primaire motorische cortex), met verbindingen met basale ganglia en cerebellum. In hun studie vonden zij dat het DCM bewijs insula in een positie tussen BA44 en twee parallelle knopen die de cerebellum/basale ganglia en motorische cortex omvatten. Heim et al., (2009) beoordeelde de afferente connectiviteit met BA45 (en ba44, indirect) in een taaltaak van fonologische/lexicale discriminatie visueel gepresenteerd.
twee aanvullende studies hebben zich gericht op het verkavelen van Broca ‘ s gebied met betrekking tot structuur en functie. De eerste studie gebruikte probabilistische tractografie en omvatte BA45. Drie gescheiden gebieden werden geïdentificeerd: BA44, BA45, en diepe operculaire gebied grenzend aan de anterior insula (Anwander et al., 2007); de tweede studie maakte gebruik van een methode gebaseerd op patronen van co-activering op verschillende verschillende cognitieve taken., In dit werk beschrijven de auteurs vijf onderverdelingen van het gebied van de Broca, twee achterste gebieden met betrekking tot fonologie (dorsaal) en ritmische sequencing (ventraal), en drie voorste gebieden met betrekking tot werkgeheugen, switching control en semantiek (Clos et al., 2013). Het is niet duidelijk hoe deze verkaveling harmoniseert met eerdere bevindingen die een dorso-ventrale differentiatie van BA44 beschrijven, waarbij in het dorsale aspect het observatiegerelateerde spiegelneuronsysteem (Molnar-Szakacs et al., 2005).
onze methode hoewel nieuw is niet volledig nieuw., Sundermann en Pfleiderer (2012) voerden een studie uit met dezelfde methodologie als onze studie. Deze auteurs, echter, niet gericht taal. In plaats daarvan richtten ze hun studie op de “inferior frontal junction” (een gebied dat inferior frontal gyrus, caudaal aspect van de middle frontal gyrus en anterior insula omvat), en zijn rol in cognitieve controle. Ze pasten meta-analytical connectivity modeling (MACM) toe op basis van de ALE-methode. Zoals wij deden in onze benadering, deden zij hun analyse die alle artikelen samenvoegen waarin de linker of rechter inferieure frontale gyri werden gerapporteerd als geactiveerd., Net als wij maakten zij ook van het activeringsgebied de onafhankelijke variabele zonder specifieke aannames met betrekking tot functionele specialisatie van het doelgebied. Alle co-activeringsgebieden werden geacht te zijn verbonden met het kerngebied (selectiecriteria voor de MACM).
het belang van het karakteriseren van het taalnetwerk van BA44 ligt in het eerder genoemde feit van multimodaliteit betrokkenheid van dit gebied over verschillende domeinen in cognitie. Broca ‘ s gebied als de kern van expressieve taal, lijkt andere vele functies te hebben., Naast de reeds genoemde functies lijkt BA44 betrokken te zijn bij verbale werkgeheugentaken (Rämä et al., 2001; Sun et al., 2005), in het bijzonder geheugen van het syntactische type (Fiebach et al., 2005; Wang et al., 2008); mirror neuron system (Manthey et al., 2003; Lawrence et al., 2006; Lotze et al., 2006); motor programing (Amunts et al., 2004); tactile imagery (Yoo et al., 2003); rekenkundige verwerking (Rickard et al., 2000); en zelfs muziekgenot (Koelsch et al., 2006)., Deze multifunctionaliteit van Broca ‘ s gebied kan gedeeltelijk worden verklaard door de anatomische onderverdeling die eerder is beschreven, maar nog steeds moeten specifieke deelgebieden op een specifieke manier met elkaar verbinden en verschillende taakgerelateerde netwerkconfiguraties produceren.de sub-anatomische differentiatiehypothese van Broca wordt ondersteund door een recent histologisch autoradiografieonderzoek dat verschillende populaties celreceptoren aantoont (Amunts et al., 2010). De belangrijkste aanname in dit werk is dat de differentiatie in celreceptor segmenten ook de functie., Er zijn zes receptortypes die Broca ‘ s in een (1) ventrale precentrale overgangsschors, (2) dorsale BA44, (3) ventrale BA44, (4) anterior BA45, (5) posterior BA45 en (6) middle frontal gyrus frontal operculum. Van belang is de bredere betekenis van Broca ‘ s gebied in dit werk, waaronder BA45 of par triangularis van het IFG.
al deze feiten zijn indicatief voor een multi-potentiele functie van BA44 die hoogstwaarschijnlijk expressie heeft in een configuratie met meerdere netwerken die de verschillende functie-uitvoer vertegenwoordigt., Daarom is een karakterisering van alle mogelijke netwerkconfiguraties raadzaam in een poging om de plasticiteit van de hersenfunctie en de mogelijke klinische effecten van de lokale laesies te begrijpen.
onze resultaten hebben belangrijke implicaties. Het kan dienen als vertrekpunt om de verschillende Broca-gerelateerde netwerken geassocieerd met diverse hersenfuncties verder te caractheriseren. Het kan ook gemakkelijk de verklaring van de complexiteit van syndromen gezien in spraak en taalstoornissen, moeilijk te reduceren tot twee of drie modules van de taal standaard model., De demonstratie van specifieke netwerken die ook specifieke cognitieve functies dienen, zoals voorgesteld in dit artikel, is belangrijk voor kruisvalidatie van andere technieken die hersenconnectiviteit aantonen; het kan ook dienen om de functionele betrokkenheid van structurele connectiviteit te bewijzen of dysproof. Bijvoorbeeld, is gebleken dat sommige onderwerpen met rechter hemisfeer dominantie voor taal, hebben verlaten boogvormige fasciculus dominantie (Dick et al.,, 2013), in welk geval, de structurele connectiviteit niet volgt neurale connectiviteit; onze methode kan ook evolueren als een hulpmiddel om een bepaalde taal lateralisatie als afzonderlijke rechts naar links connectiviteit worden aangetoond in toekomstig onderzoek op dit gebied.
beperkingen
veel meer artikelen hebben BA44-activering in taaltaken beschreven, maar ze zijn niet opgenomen in de brainmap.org database., Om de database in te voeren, moeten de studieresultaten de activering rapporteren in standaard ruimtecoördinaten (Mni of Talairach), die handmatig in de database moeten worden ingevoerd en vervolgens moeten worden goedgekeurd door het team dat de database leidt. brainmap.org project. Ondanks deze beperking schatten de auteurs dat het aantal studies/deelnemers/experimenten dat de pooling-data binnenkomt groot is en de state of the art publicaties in fMRI van taal weerspiegelt.
een andere potentiële beperking die voortvloeit uit de voorbewerking is de verkeerde toewijzing van activering in gebieden waar twee verschillende kwabben of structuren aan elkaar grenzen., Een deel van de voorbewerking van de gegevens bestaat uit het gladstrijken van de activering. In smoothing, voxels met een lager signaal dan de buren worden verhoogd voor minder luidruchtige presentatie. Dus, het algoritme behandelt alle buren als een continuüm. Deze procedure verklaart wat hoogstwaarschijnlijk een valse activering van de culmen van het cerebellum is, aangezien het gladstrijken van de ware activering verkregen in de spoelvormige gyrus de culmen nadert. Een soortgelijk effect verklaart de roep van activering in de rechter anterior cingulate gyrus, en de linker temporale superior gyrus (gebied 22 in cluster 1)., Deze activering is hoogstwaarschijnlijk te wijten aan de verlenging van het gladmaken van het aangrenzende frontale operculum.
twee extra beperkingen moeten worden blootgesteld. Ondanks de Duidelijke betrokkenheid van BA45 in de taal die is aangetoond door fMRI studies hierboven, wordt dit gebied niet vermeld in het ALE automatic rapport. Cluster 1 (Tabel 1) toont de activering in alle gebieden rond BA45 (d.w.z. 44, 47, 6 en 9)., Daarom lijkt het erop dat het algoritme een “blok” aanneemt inclusief BA45 (Pars Tringularis) binnen BA44, aangezien de activering openlijk is in dat gebied volgens het weergegeven beeld (figuur 1, rechterpaneel). Een soortgelijke situatie kan het gebrek aan verslag van BA21 verklaren. Deze technologische beperkingen zijn verre van geschikt om door de auteurs te worden gewijzigd. Op zich hebben zij echter geen invloed op de statistische analyse of resultaten.
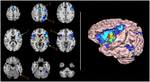
figuur 1. Functionele connectiviteit kaart van BA44 door Meta-analytische connectiviteit modellering., (Links) transversale dalende sneden van de hersenen MRI-sjabloon. Linkerhersenhelft verschijnt aan de rechterkant (radiologische conventie). Clusters van activering zijn kleur gecodeerd voor statistische significantie van donkerblauw (laagste) tot rood (hoogste). Clusternummers van het rapport van de automatische activeringswaarschuwingsschatting (ALE) worden geassocieerd met de belangrijkste beeldclusters. Pijlen wijzen Ongeveer naar hun isocenters. Binnen het gele ovaal, komt cluster 7 overeen met de linker thalamus, met mediale lokalisatie, en cluster 8, lenticulaire kern, met laterale en rostrale positie., De cerebellaire activering in het midden van de onderste rij maakt deel uit van cluster 5. Het wordt waarschijnlijk verklaard door het gladmakende effect van de aangrenzende activering van de linker spoelvormige gyrus. (Rechts) 3D volumetrische weergave van de hersenen met activering op het oppervlak van de linkerhersenhelft. Rode kleurzone identificeert BA44. Diepe en middellijn activeringen worden niet getoond.,
andere beperkingen zijn bewuste beperkingen van de studie die bedoeld zijn om het effect van verstorende factoren te vermijden waarvan bekend is dat ze van invloed zijn op de taal lateralisatie (righ-handed normale proefpersonen, taaltype: occidental, en leeftijd: 20-60). Het onderzoek naar de wisselwerking tussen deze variabelen en het netwerk van Broca is de moeite waard om in toekomstig onderzoek te worden aangepakt.
de komst van een grote database maakt het mogelijk om informatie betrouwbaar te aggregeren onder een bepaalde variabele. Het zou ons in staat stellen om betrouwbare informatie te verkrijgen met een hoog vermogen tot veralgemening., Bovendien maakt het samenvoegen van gegevens in de methode die we hier voorstellen het mogelijk om gebieden van co-activering te demonstreren over onderwerpen en over taken die ofwel een specifieke functie of een groep Functies met betrekking tot een specifiek domein ondersteunen. In het bijzonder kan elke specifieke taak (fonologie, semantiek, begrip, enz.) slechts een paar van de modules gebruiken. De weergave van hoge specifieke netwerken die specifieke functies vervullen, kan van belang zijn in de klinische praktijk., Bijvoorbeeld, kon het netwerken met betrekking tot lateralisatie van taal identificeren, en vandaar om in neurochirurgische planning bij patiënten niet geschikt voor taak-afhankelijke fMRI of Wada tests te helpen. In het algemeen, onze methode kan aantonen dat de mogelijke of potentiële connectiviteit van het netwerk voor dat cognitieve domein verbeteren van het begrip van de hersenfunctie., We zouden bijvoorbeeld alle verschillende “configuraties” waarin BA44 deelneemt kunnen beoordelen om de maximale connectiviteit te onthullen of, op kleinere schaal, naar differentiatie van dergelijke configuraties kunnen zoeken om te verduidelijken hoe BA44 betrokken is bij verschillende functies.
conclusie
we hebben de toepassing van een pooling data methode gedemonstreerd om het netwerk van BA44 gerelateerd aan taal in het algemeen weer te geven. De gevonden clusters van activering zijn in overeenstemming met eerdere klinische en neuroimaging studies, hoewel de laatste schaars zijn., Om de hersenfunctie te verklaren, zal de beschrijving van netwerken in de immidiate toekomst een grotere impact hebben dan de beschrijving van hersenmodules in het verleden zou hebben gehad. Beter begrip van de hersenconnectiviteit zal noodzakelijkerwijze helpen om hersenfuncties beter te begrijpen.
belangenconflict verklaring
De auteurs verklaren dat het onderzoek werd uitgevoerd zonder enige commerciële of financiële relatie die als een potentieel belangenconflict kon worden opgevat.,
voetnoten
- ^http://brainmap.org
- ^http://brainmap.org
- ^http://ric.uthscsa.edu/mango/
Benson, F., and ardila, A. (1996). Afasie: Een Klinisch Perspectief. Oxford: Oxford University Press.
Google Scholar
Broca, P. (1861). Opmerkingen over de zetel van de faculteit articulate language; gevolgd door een waarneming van aphemie. Stier. Soc. Antropol. 2, 330–357.
Google Scholar
Brodmann, K. (1909)., Vergelijkende lokalisatietheorie van de hersenschors gepresenteerd in zijn principes op basis van celconstructie. Leipzig: Barth-Verlag.