Innledning
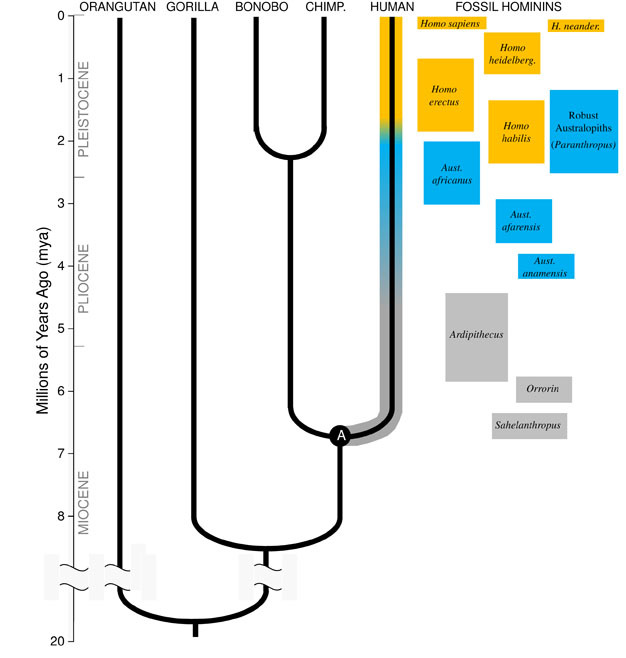
Darwin ‘ s god innsikt, og den samlende prinsipp i biologi i dag, er at alle arter er knyttet til hverandre som søstre, fettere og kusiner, og fjerne slektninger i en stor familie tre av livet. Implikasjonene er fantastisk; hvis vi kunne reise langt nok tilbake i tid, ville vi finne felles aner mellom oss selv og alle andre levende organisme, fra pinnsvin å flamingoer å kaktuser., Vår umiddelbare evolusjonære familie består av hominoids, gruppen av primater som inkluderer «mindre aper» (siamangs og gibbons) så vel som den «store aper» (sjimpanser, bonobos, gorillaer og orangutanger). Blant de store apene, våre nærmeste slektninger er sjimpanser og bonobos (Figur 1). Den fossil-posten, sammen med studier av menneske og ape DNA, tyder på at mennesker delte en felles stamfar med sjimpanser og bonobos en gang rundt 6 millioner år siden (mya)., Vi begynner denne diskusjonen av våre artens utvikling i Afrika, i nærheten slutten av den geologiske perioden som er kjent som Miocene, like før vår ætt skilte seg fra det som sjimpanser og bonobos.
Miocene Opprinnelsen til Hominin Avstamning
for å forstå utviklingen av enhver art, må vi først etablere sine forfedres state: hva slags dyr gjorde det utvikle seg? For vår ætt, dette krever at vi prøver og rekonstruere den Siste Felles Stamfar av mennesker og sjimpanser (merket med «A» i Figur 1). Human-Sjimpanse Siste Felles Stamfar (HC-LCA) er arter som hominin avstamning og sjimpanse & bonobo ætt skilte seg., Hominins er arter på vår gren av hominoid tre etter splitt med sjimpanse & bonobo linje, inkludert alle de utdødde arter og viktige sidegreiner (Figur 1).
Det var et stort mangfold av ape arter i Miocene, med dusinvis av arter som er kjent fra fossile ta opp hele Afrika, Europa og Asia. Disse artene varierte i deres anatomi og økologi, og det er ikke klart hvilke, om noen, av de fossile arter oppdaget så langt representerer HC-LCA (Kunimatsu et al. 2007; Unge og MacLatchy, 2004)., Likevel vet vi fra fossile og komparativ bevis for at det var mye mer lik lever aper enn til levende mennesker. HC-LCA ville ha hatt en ape-størrelse hjernen og kroppen, med relativt lange armer og fingre og ta tak i foten som gjorde det mulig å grovfôr i trærne. Hjørnetann tenner var trolig stor og skarp, som vi har sett i flere Miocene hominoids. Videre hjørnetenner var trolig seksuelt dimorphic, med menn som har mye større hjørnetenner enn kvinner, som vi har sett blant de levende store aper og Miocene fossiler., Som lever aper ville det ha gått quadrupedally (på alle fire) når du er på bakken, og dens diett ville ha besto nesten utelukkende av planteføde, først og fremst frukt og blader.
Tidlig Hominins
Endres fra en ape-lignende anatomi er merkbar i hominoid fossiler fra slutten av Miocene i Afrika. Noen hominoid arter fra denne perioden viser trekk som er typiske for mennesker, men er ikke sett i andre lever aper, ledende paleoanthropologists å antyde at disse fossiler representerer tidlige medlemmer av hominin avstamning., Den første menneskelignende trekk til å vises i hominin fossilt posten er tobent gange og mindre, sløv hjørnetenner.
De eldste hominins tiden kjent er Sahelanthropus tchadensis fra Tsjad (Brunet et al. 2005) og Orrorin tugenensis fra Kenya (Senut et al. 2001). Sahelanthropus, datert til mellom 6 og 7 mya, er kjent fra en stor grad komplett skallen og noen andre fragmentariske rester. Hjernen størrelse, 360cc, er innenfor rekkevidde sett i sjimpanser, og skallen har en massiv panne ridge, like i tykkelse for å mannlige gorillaer (Brunet et al. 2005)., Men, posisjon og orientering av foramen magnum, hullet i bunnen av skallen der ryggmargen går, tyder på at Sahelanthropus sto og gikk bipedally, med sin ryggsøylen holdt vertikalt som i moderne mennesker i stedet for horisontalt som i aper og andre firføtte (Zollikofer et al. 2005). Orrorin er først og fremst kjent fra postcranial fossiler, inkludert en delvis femur. Den proksimale del av femur viser likheter med de moderne mennesker, noe som tyder på at arten ble bipedal (Pickford et al. 2002)., Ingen hodeskaller av Orrorin har blitt gjenopprettet, og så dens cranial morfologi og hjernen størrelse er usikre. I begge Orrorin og Sahelanthropus hjørnetann tenner på menn er større og skarpere enn hos moderne mennesker, men er liten og butt i forhold til hjørnetenner av mannlige aper. Dette tyder på at canine seksuell dimorphism — og i forlengelsen av dette, konkurranse blant menn for parring tilgang til kvinner — ble redusert i disse tidlige hominins i forhold til de store apene.
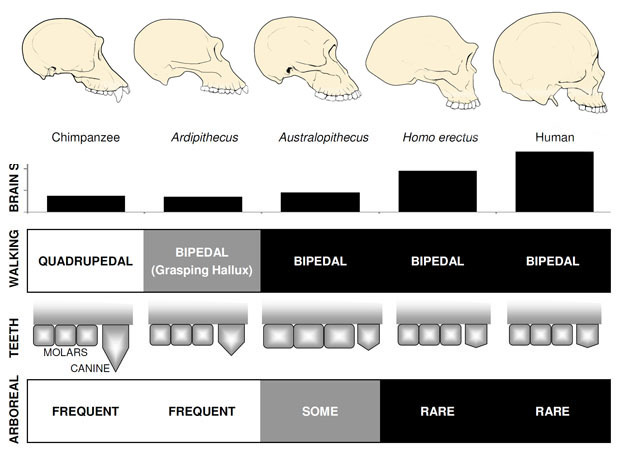
desidert best kjent tidlig hominin er Ardipithecus ramidus, en 4.,4 millioner år gamle arter fra Etiopia, som er kjent fra en nesten komplett skjelett samt en rekke andre dental og skjelettrester (White et al. 2009). Ar. ramidus og en eldre, beslektede arter kjent fra fragmentariske rester, Ar. kadabba (5.8–5.2 mya), har redusert hjørnetenner som ligner på de av Orrorin og Sahelanthropus. Skallen til Ar. ramidus er heller apelignende og bredt lik som Sahelanthropus, med en liten sjimpanse-størrelse hjernen på 300–350cc (Figur 2). Den Ardipithecus postcranial skjelettet er spennende., Selv om svært fragmentert, bekkenet utvinnes avslører en morfologi ganske forskjellig fra levende aper, med en kortere, mer bowl-lignende form som sterkt antyder Ardipithecus gikk bipedally; dette er i samsvar med foramen magnum posisjon, noe som tyder på en oppreist kroppsholdning. Imidlertid, sin lange forelimbs og fingre og sin avvikende, fatte første tå (hallux) foreslår Ardipithecus tilbrakte mye av sin tid i trærne. Det generelle inntrykket er av et stor grad trefylt arter som gikk bipedally når det våget seg til bakken.,
Australopithecus
Rundt 4mya finner vi de tidligste medlemmer av slekten Australopithecus, hominins som var flinke terrestriske bipeds, men fortsatte å bruke trær for mat og beskyttelse. De første prøvene av Australopithecus ble oppdaget i Sør-Afrika i 1924 (Dart, 1925), og forskning innsats over de påfølgende åtte tiår har produsert hundrevis av fossiler fra flere arter på alle områder på tvers av Øst-og Sørlige Afrika., Vi vet nå at Australopithecus var en svært vellykket slekten som vedvarte i nesten tre millioner år (Figur 1).
Den mest kjente Australopithecus arter A. afarensis (3.6–2.9 mya) fra Øst-Afrika og A. africanus (3.2–2.0 mya) fra Sør-Afrika., Bekkenet og nedre lemmer av disse artene indikerer klart og tydelig at de var fullt bipedal: bekken er kort og bolle-formet, noe som gluteal musklene rundt til side av kroppen, som i moderne mennesker, for stammen stabilisering under bipedalism, og den første tå er på linje med de andre tærne (Ward, 2002; Harcourt-Smith og Aiello, 2004). Den Australopithecus foten kan også ha hatt en menneskelignende arch, basert på analyse av metatarsals og det fossile Laetoli avtrykk (Ward et al. 2011)., Likevel, i forhold til moderne mennesker, underarmene var lange og fingre og tær var lang og noe buet, noe som tyder på at Australopithecus regelmessig brukes trær til fôr og kanskje som en tilflukt fra rovdyr på natten. Dette blandet terrestriske & trefylt strategi ville ha servert disse artene også i blandet skog og savanne miljøer de er bebodd.
Hjernen størrelse i Australopithecus varierte mellom 390 og 515cc, lik sjimpanser og gorillaer (Falk et al., 2000), noe som tyder på kognitive evner var i store trekk lik lever aper (Figur 2). Body størrelse i Australopithecus var ganske liten og seksuelt dimorphic, om 30kg for kvinner og 40kg for menn (McHenry, 1992). Dette nivået av dimorphism er ikke reflektert i hjørnetenner, som var liten, butt, og bare som i tidligere hominins.
i Motsetning til den hjørnetenner, molar tenner i Australopithecus var mye større enn de tidligere hominins, og hadde tykkere emalje. Dette tyder på kostholdet, inkludert vanskelig, lav kvalitet planteføde som kreves kraftig tygging til prosessen., En undergruppe av Australopithecus, kjent som «robust» australopiths (ofte merket med et eget slekten Paranthropus) på grunn av deres enorme tenner og tygge muskler, tok denne tilpasningen til det ekstreme. De fleste Australopithecus arter ble utryddet av 2 mya, men noen robuste former vedvarte inntil om lag 1,2 mya i Øst-og Sør-Afrika.
Slekten Homo
De eldste fossiler av vår egen slekten Homo, er funnet i Øst-Afrika, og datert til 2.3 mya (Kimbel et al. 1997)., Disse tidlige prøvene er lik i hjernen og kroppen størrelse til Australopithecus, men viser forskjeller i deres molar tennene, noe som tyder på en endring i kostholdet. Faktisk, med minst 1.8 mya, tidlige medlemmer av vår slekten var med primitive stein verktøy til å slakte dyret er slaktet, og legger til energi-rik kjøtt og bein marg i sine plantebasert diett.
De eldste medlem av slekten Homo, H. habilis (2.3–1.4 mya) er funnet i Øst-Afrika, og er forbundet med slaktede dyr bein og enkel stein verktøy (Blumenschine et al. 2003). Det er mer formidabel og utbredt etterkommer, H., erectus, er funnet i hele Afrika og Eurasia, og vedvarte fra 1,9 mya til 100 kya, og kanskje enda senere (Anton, 2003). Som moderne mennesker, H. erectus manglet forelimb tilpasninger for klatring sett i Australopithecus (Figur 2). Sin globale ekspansjon foreslår H. erectus var økologisk fleksibel, med den kognitive kapasiteten til å tilpasse seg og trives i svært ulike miljøer. Ikke overraskende, det er med H. erectus at vi begynner å se en stor økning i hjernens størrelse, opp til 1,250 cc for senere Asiatiske prøver (Anton, 2003). Molar størrelsen er redusert i H., erectus i forhold til Australopithecus, noe som gjenspeiler sin mykere, fyldigere kosthold.
Rundt 700 kya, og kanskje tidligere, H. erectus i Afrika ga opphav til H. heidelbergensis, en art veldig mye som oss i form av kroppens proporsjoner, dental tilpasninger, og kognitive evnen (Rightmire, 2009). H. heidelbergensis, ofte referert til som en «arkaisk» Homo sapiens, var en aktiv big-game hunter, produsert sofistikert Levallois stil verktøy, og med minst 400 kya hadde lært å kontrollere brann (Roebroeks og Villa, 2011). Neandertalerne (H., neanderthalensis), tilpasset kulde hominins med stout physiques, komplekse atferd, og hjernen lignende i størrelse til oss, er antatt å ha utviklet seg fra H. heidelbergensis populasjoner i Europa med minst 250 kya (Rightmire, 2008; Hublin, 2009).
Fossil og DNA-bevis foreslå vår egen art, H. sapiens utviklet seg i Afrika 200 kya (Relethford, 2008; Rightmire, 2009), trolig fra H. heidelbergensis. Den økte atferdsvansker raffinement av H., sapiens, som indikert av våre store hjerner (1,400 cc) og arkeologiske bevis for en mer omfattende verktøysett og smart jakt teknikker, lov vår arter til å blomstre og vokse på det Afrikanske kontinentet. Ved 100kya, våre arter sølt i Eurasia, slutt å utvide over hele verden og inn i Australia og Amerika (DiGiorgio et al. 2009). Langs veien våre arter fortrenges andre hominins de møtte, inkludert Neandertalerne i Europa og lignende former i Asia. (Merk at ikke alle er enige i denne tolkningen av data, se Tryon og Bailey)., Studier av gammelt DNA ekstrahert fra Neandertaler-fossiler antyder våre arter kan ha tidvis interbred med dem (Green et al., 2010). Vår økende global impact fortsetter i dag, som kulturelle innovasjoner som for eksempel landbruk og urbanisering forme landskapet og arter som er rundt oss.
Oppsummering
utviklingen av våre arter fra en ape-lignende Miocene stamfar var en kompleks prosess. Vår avstamning er full av sidegreiner og evolusjonære blindveier, med arter som robust australopiths som vedvarte i over en million år før forsvinne., Noen menneskelige trekk, som bipedalism, utviklet seg svært tidlig, mens andre, som for eksempel store hjerner, ikke utvikle seg til relativt nylig. Fremdeles andre egenskaper, som molar størrelse, utviklet seg i en retning bare for å bli presset tilbake senere ved å endre økologisk påvirkning. Snarere enn en kraftig skipet kartlegging en rett kurs mot noen forhåndsbestemt mål, utvikling av vår ætt, ja, av alle arter’ avstamning — passer det bildet av en livbåt kastet omkring av skiftende hav av miljøendringer, genetisk flaks, og geologiske sjanse. Man lurer på hvor den neste seks millioner år kan ta oss.