소개
다윈의 훌륭한 통찰력 및 통합 생물학의 원리로 오늘날에는 모든 종은 관련된 하나의 또 다른 자매 같은,사촌,먼 친척에 광대한 가족 생명의 나무. 영향은 아름다운 경우 우리가 충분히 시간에서,우리가 찾을 것이 일반적인 조상들이 우리 자신과 다른 모든 생명체에서 고슴도치를 홍학을 선인장., 우리의 즉각적인 진화 가족의 구성 hominoids,그룹의 영장류가 포함된”적은 원숭이”(siamangs 및 기븐스)뿐만 아니라”좋은”원숭이(침팬지,보노보,고릴라,오랑우탄). 유인원 중 가장 가까운 친척은 침팬지와 보노보입니다(그림 1). 화석 기록과 함께,연구 결과는 인간과 원숭이 DNA 나타냅니다 인간은 공통 조상으로 침팬지 및 보노보가 6 백만 년 전(mya)., 우리가 시작이 토론 우리의 종의’진화에서 아프리카 근처의 끝 지질학적 기간으로 알려진 중신세,단지기 전에 우리의 혈통에서 분기의 것 및 침팬지죠.
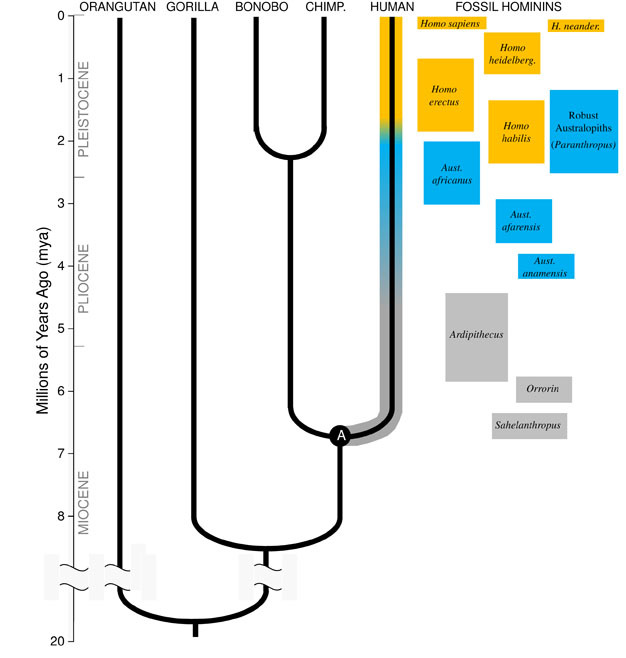
메탄은 기원의 완전하게 채우는 방식이 Lineage
을 이해하기 위해서는 진화의 종,우리는 먼저 설정 그 조상의 상태:의 어떤 종류의 동물은 그것에서 진화? 에 대한 우리의 계보,이 필요로 하는 우리는 시도하고 재구성 하는 마지막 일반적인 조상의 인간 및 침팬지(표시된”A”에서 그림 1). 인간 침팬지의 마지막 일반적인 조상(HC-LCA)은 종에서는 완전하게 채우는 방식이 혈통 및 침팬지&보노보통 갈라집니다., Hominins 은 종에 우리의 지점의 hominoid 트리 분할 후으로 침팬지&보노보 라인에 포함하여 모든의 멸종과 진화 가지 측면(그림 1).
이 있었 다양한 원숭이 종에서 중신세,수십 종의에서 알려진 화석 기록에서 아프리카,유럽 및 아시아. 이 종들은 해부학과 생태학에서 다양했으며,지금까지 발견 된 화석 종 중 어느 것이 HC-LCA 를 나타내는 지 명확하지 않다(Kunimatsu et al. 2007;젊고 MacLatchy,2004)., 그럼에도 불구하고,우리가 알고있는 화석연하고 비교는 증거 그것은 훨씬 더 많은 비슷하게 생활하는 것보다 원숭이 살아있는 인간. HC-LCA 했을 것이 원숭이-크기의 두뇌와 몸으로,상대적으로 긴 팔과 손가락과 파악 발는 그것을 허용 사료에 나무입니다. 송곳니 치아는 아마도 여러 미오세 호미 노이드에서 볼 수 있듯이 크고 날카 로웠을 것입니다. 또한,송곳은 아마도 성적으로 동종 이형을 가진 남자 하는 데는 훨씬 더 큰 개 여성보다,본 사이 살아있는 유인원과 메탄은 화석을 볼 수 있습니다., 같은 생활을 원숭이가 걸어 quadrupedally(발로)지상에 있을 때,그것의 다이어트로 구성되어 있의 거의 전적으로 식물성 음식,주로 과일과 나뭇잎.
초기 Hominins
원숭이와 같은 해부학의 변화는 아프리카의 후기 Miocene 의 hominoid 화석에서 식별 할 수있다. 일부 hominoid 에서 종이 이 기간에 전시는 특성을의 전형적인 인간지에 보이지 않는 다른 살아있는 원숭이,선도 paleoanthropologists 을 추론하는 이 화석들 대표 초기 구성원의 완전하게 채우는 방식이랍니다., 호미닌 화석 기록에 나타나는 최초의 인간과 같은 특성은 두발 보행과 더 작고 뭉툭한 송곳니입니다.
현재 알려진 가장 오래된 호미닌은 차드의 Sahelanthropus tchadensis(Brunet et al. 2005)및 케냐의 Orrorin tugenensis(Senut et al. 2001). 6 에서 7mya 사이의 sahelanthropus 는 크게 완전한 두개골과 다른 단편적인 유적으로 알려져 있습니다. 그 뇌 크기,360cc,는 범위 내에서 본 침팬지,그리고 이러한 배경에는 대규모 이마 리지,비슷한에 두께하는 남성의 고릴라(Brunet et al. 2005)., 그러나,위치 및 방향의 구멍 매그넘는 구멍의 기초에 두개골을 통해 척추 전달,건 Sahelanthropus 서서하고 걸어 bipedally,그 척추 수직으로 개최되으로 현대적인 인간이 아닌 수평으로서 원숭이고 다른 네발(Zollikofer et al. 2005). Orrorin 은 주로 부분 대퇴골을 포함한 후두 화석에서 알려져 있습니다. 대퇴골의 근위 부분은 현대 인간의 것과 유사성을 보여 주며,종은 두발 형 이었음을 시사한다(Pickford et al. 2002)., Orrorin 의 두개골은 회복되지 않았으므로 두개골 형태와 뇌 크기는 불확실합니다. Orrorin 과 Sahelanthropus 모두에서 수컷의 송곳니 이빨은 현대인보다 크고 뾰족하지만 수컷 유인원의 송곳니에 비해 작고 둔합니다. 이는 개를 성적이 형태 및으로 확장을,경쟁에 남성들 사이에 결합에 대한 액세스를 여성이었 감소에서 이러한 초기 hominins 에 비해 훌륭한렵습니다.
지금까지 가장 잘 알려진 초기 hominin 은 Ardipithecus ramidus,a4 입니다.,거의 완전한 골격뿐만 아니라 수많은 다른 치과 및 골격 유적에서 알려진 에티오피아에서 4 백만년 된 종(White et al. 2009). 아칸소. ramidus 및 이전,단편적인 유물에서 알려진 관련 종,Ar. kadabba(5.8–5.2mya),Orrorin 과 Sahelanthropus 의 개과 유사한 개과가 감소했다. 아칸소의 두개골. ramidus 는 300-350cc 의 작은 침팬지 크기의 뇌를 가진 sahelanthropus 와 다소 원숭이 같고 대체로 유사합니다(그림 2). Ardipithecus postcranial 골격은 흥미 롭습니다., 비록 심하게 조각,골반 회복을 보여준에서 상당히 다른 형태의 생 원숭이,짧은,더 그릇 같은 모양이 강하게 제안 Ardipithecus 걸 bipedally;이것은 일관적으로 구멍 매그넘치는 수직 자세입니다. 그러나,그것의 오래 앞다리와 손가락과 발산,장악은 첫째 발가락(정)건 Ardipithecus 하는 데 많은 시간의 나무에 있습니다. 전반적인 인상은 땅에 모험을 할 때마다 두발로 걷는 크게 수목 종입니다.,
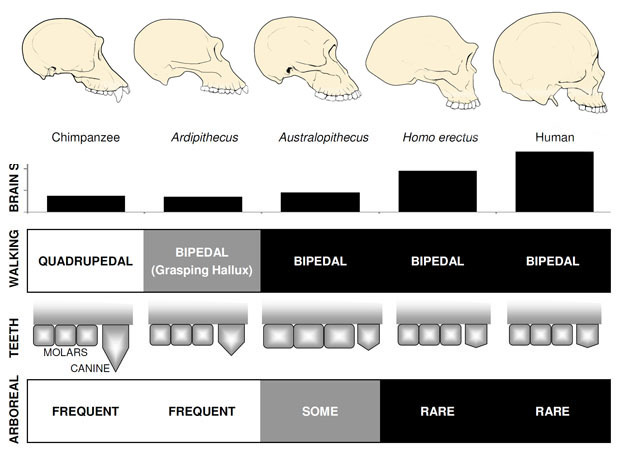
오
주 4mya 우리가 찾을 초기 구성원 속의 오,hominins 는 숙련 지상파 사는 두발 승 를 십만 계속 사용하고 나무를 음식과 보호합니다. 첫 번째 표본의 오서 발견되었습 남아프리카 공화국에서는 1924 년(Dart,1925)및 연구 활동을 통해 이후의 여덟 수십 년 동안 생산의 수백 화석에서 여러 가지 종의 사이트 모두에서 동쪽과 남아프리카., 우리가 알고 있는 오 매우 성공적이 속하는 지속되는 거의 삼백만 년습니다(그림 1).
가장 잘 알려진 오 종 A. 따라(는 3.6 2.9mya)동쪽에서 아프리카와 아프리카누스(3.2–2.0mya)에서 남아프리카., 골반과 더 낮은 사지의 이러한 종을 명확하게 나타나는 것들이 완전히 두발:골반은 짧고 그릇 모양이를 데리고,둔부 근육을 측면의 몸으로,현대에 인간들,트렁크하는 동안 안정 직립보행의 부수적,그리고 첫 번째 발가락이 라인에서 다른 발가락(병동,2002;하 커트-스미스 및 Aiello,2004). 오스트랄로 피테쿠스 발은 중족골과 화석화 된 라에 톨리 발자국(Ward et al. 2011)., 그럼에도 불구하고,비교하여 현대의 인간,팔던 길과 손가락과 발가락이 오랫동안 다소,곡선이 제안하는 오는 정기적으로 사용되는 나무하는 꼴을 주고 아마도 피난처로서 육식에서 밤입니다. 이 혼합된 지구&나무 전략가 제공되는 이러한 종에서 잘 혼합된 삼림과 사바나에 환경은 그들이 거주하고 있습니다.
오스트랄로 피테쿠스의 뇌 크기는 침팬지와 고릴라와 비슷한 390~515cc 범위였다(Falk et al., 2000),인지 능력이 살아있는 유인원과 대체로 유사하다는 것을 시사한다(그림 2). 오스트랄로 피테쿠스의 신체 크기는 다소 작고 성적으로 이형성이었으며 여성의 경우 약 30kg,남성의 경우 40kg 이었다(McHenry,1992). 이 이형성의 수준은 이전 호미닌에서와 같이 작고 둔하며 단형 인 송곳니에는 반영되지 않습니다.
송곳니와는 달리,오스트랄로 피테쿠스의 어금니는 이전의 호미닌보다 훨씬 컸으며 더 두꺼운 에나멜을 가지고있었습니다. 이것은 그들의 식단이 가공하기 위해 강력한 씹기가 필요한 단단하고 품질이 낮은 식물성 식품을 포함했음을 시사합니다., 하위의 오으로 알려진”강력한”australopiths(종종 레이블이 붙여 별도의 속 Paranthropus)기 때문에 그들의 엄청난 이빨고 씹는 근육,이 적응을 극단적이다. 대부분의 오 종의 멸종되었으로 2mya 하지만,일부 강력한 형태로 유지 될 때까지 약 1.2mya 에서 동쪽과 남아프리카.
속 호모
우리 자신의 속의 가장 초기의 화석 인 호모(Homo)는 동 아프리카에서 발견되며 2.3mya 로 거슬러 올라간다(Kimbel et al. 1997)., 이 초기 표본은 오스트랄로 피테쿠스와 뇌 및 신체 크기가 비슷하지만 어금니의 차이를 보여서식이 요법의 변화를 시사합니다. 실제로,적어도 1.8mya,초기 구성원이 우리의 속에 사용하던 원시 돌 도구를 정육점이 동물의 사체,에너지 추가 풍부한 고기와 골수를 그들의 식물 기반의 식단입니다.
가장 오래된 회원의 종,H. 커(2.3–1.4mya)에서 발견과 아프리카 동부지역과 연결된 도살 동물 뼈와 간단한 돌 도구(Blumenschine et al. 2003). 그것의 더 강력하고 널리 퍼진 후손,H., 에렉투스는 아프리카와 유라시아 전역에서 발견되며 1.9mya 에서 100kya 까지,그리고 아마도 나중에(Anton,2003)지속되었습니다. 현대 인간과 마찬가지로 H.erectus 는 오스트랄로 피테쿠스에서 볼 수있는 등반을위한 앞다리 적응이 부족했습니다(그림 2). 그것의 세계적인 확장은 H.erectus 가 크게 다른 환경에서 적응하고 번성 할 수있는인지 능력과 함께 생태 학적으로 유연하다는 것을 암시합니다. 당연히 H.erectus 와 함께 우리는 후기 아시아 표본에 대해 최대 1,250cc 의 뇌 크기가 크게 증가하기 시작합니다(Anton,2003). 몰 크기는 H 에서 감소된다., 쪽에 상대적인 오 반영하여 부드럽고,풍부한 다이어트입니다.
700kya,그리고 아마도 이전에,H. 쪽 아프리카에서 상승했 H.heidelbergensis,종이처럼 매우 우리의 관점에서 신체 비율,치과 적응과 인지능력(Rightmire,2009). H.heidelbergensis,수시로 불리는”오래된”호모 사피엔스 적극적으로 큰 게임 사냥꾼,생산 르발루아 정교한 스타일의 도구로 이상 400kya 배웠었을 제어하는 불(Roebroeks 및 빌라,2011). 네안데르탈 인(H., neanderthalensis),냉 적응 hominins 뚱뚱한 체격에,복잡한 행동과 두뇌와 유사한 크기로,우리는 생각에서 진화 H.heidelbergensis 인구는 유럽에서 최소 250kya(Rightmire,2008;Hublin,2009).
화석하고 DNA 증거는 건 우리 자신의 종,h,진화에서 아프리카 200kya(Relethford,2008;Rightmire,2009),에서 아마 H.heidelbergensis. H. 의 증가 된 행동 정교함, 사피엔스로 표시하여 우리의 큰 뇌(1,400cc)및 고고 학적 증거의 광범위한 도구를 설정하고 영리한 사냥 기술,우리의 종이 번창하고 성장한 아프리카 대륙에 있습니다. 100kya 에 의해 우리 종은 유라시아로 유출되어 결국 전 세계에 걸쳐 호주와 아메리카 대륙으로 확장되었습니다(DiGiorgio et al. 2009). 길을 따라 우리의 종의 집 다른 hominins 그들이 발생하여 유럽에서 네안데르탈 인와 비슷한 양식에서 아시아입니다. (데이터의이 해석에 모두 동의하지는 않는다는 점에 유의하십시오,Tryon 과 Bailey 참조)., 네안데르탈 인 화석에서 추출한 고대 DNA 에 대한 연구는 우리 종들이 때때로 그들과 교배했을 수도 있음을 시사한다(Green et al., 2010). 우리 증가하고 글로벌 영향을 계속한 오늘날,문화 등 혁신적인 농업 및 도시화의 모양 풍경과 종 우리 주위에 있습니다.
요약
원숭이와 같은 미오세 조상에서 우리 종의 진화는 복잡한 과정이었습니다. 우리의 혈통은 전체의 측면을 가지고 진화의 죽은 끝으로,같은 종 강력한 australopiths 유지 만 이상의 년 동안하기 전에 사라져 가고있다., 이족주의와 같은 일부 인간의 특성은 매우 일찍 진화 한 반면,큰 두뇌와 같은 다른 특성은 비교적 최근까지 진화하지 않았습니다. 아직도 다른 특성을 같이,몰 크기,진화에서 한 방향으로 밀리는 것을 나중에 다시 변경하여 생태 압력이있다. 보다는 강력한 선박을 차트 똑바른 과정을 어떤 방향으로 미리 결정된 목적지,진화의 우리의 혈통에—실제로,어떤 종’혈통에 맞는 이미지의 구명보트를 빼앗에 대한 변화에 의해 바다의 환경 변화,유전자,행운 및 지질학적 기회입니다. 하나는 앞으로 6 백만 년이 우리를 어디로 데려 갈지 궁금해합니다.