はじめに
ダーウィンの偉大な洞察、そして今日の生物学の統一原理は、すべての種が、生命の広大な家系図の中で姉妹、いとこ、そして遠い親族のように互いに関連しているということです。 私たちが時間内に十分に戻って旅行することができれば、私たちはヤマアラシからフラミンゴ、サボテンまで、自分自身と他のすべての生きている生物の間に共通の祖先を見つけるでしょう。, 私たちの直接の進化の家族は、人類似、”小類人猿”(siamangsとテナガザル)だけでなく、”大類人猿”(チンパンジー、ボノボ、ゴリラ、オランウータン)を含む霊長類のグループで 大類人猿の中で、私たちの最も近い親戚はチンパンジーとボノボです(図1)。 化石記録は、ヒトおよび類人猿のDNAの研究とともに、ヒトが約6万年前(mya)にチンパンジーおよびボノボと共通の祖先を共有していたことを示している。, 私たちは、私たちの系統がチンパンジーやボノボの系統から分岐する直前に、中新世として知られている地質学的期間の終わり近くに、アフリカでの私たちの種の進化についてのこの議論を始めます。
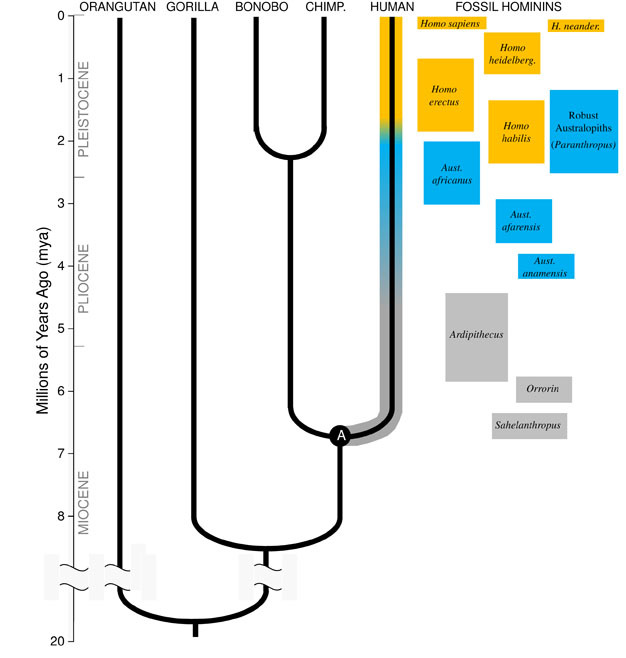
ヒト系の中新世の起源
あらゆる種の進化を理解するためには、まずその祖先の状態を確立する必要があります。 私たちの系統にとっては、ヒトとチンパンジーの最後の共通祖先(図1で”A”と記されています)を再構築しようとする必要があります。 ヒト-チンパンジー最後の共通祖先(HC-LCA)は、ヒト系とチンパンジー&ボノボ系が分岐した種である。, ホミニンは、チンパンジー&ボノボラインとの分割後のホミノイドツリーの私たちの枝の種であり、絶滅した種と進化の側枝のすべてを含む(図1)。
中新世には類人猿の種が非常に多様であり、アフリカ、ヨーロッパ、アジアの化石記録から数十の種が知られていました。 これらの種はその解剖学的および生態学的に多様であり、これまでに発見された化石種のうち、HC-LCAを表すものがどれであるかは明らかではない(Kunimatsu et al. 2007年、ヤング-アンド-マクラッチー、2004年)。, しかし、私たちは知らの化石および比較の証拠したことによるものであったり生活型類人猿以上に生活のは人です。 HC-LCAは比較的長い腕と指を持ち、木の中で餌を食べることができる掴み足を持つ類人猿の大きさの脳と体を持っていたでしょう。 いくつかの中新世の人類型に見られるように、犬の歯はおそらく大きく、鋭かった。 さらに、犬歯はおそらく性的に二形であり、雄は雌よりもはるかに大きな犬歯を有し、生きている大類人猿および中新世の化石の間で見られる。, 生きている類人猿のように、それは地面にいるときに四足歩行していたでしょう、そしてその食事はほぼ完全に植物性食品、主に果物と葉で構成されていたでしょう。
初期のヒト類
猿のような解剖学からの変化は、アフリカの後期中新世からのヒト類の化石で識別可能である。 この時期のいくつかの人類似種は、人間に典型的な特徴を示すが、他の生きている類人猿には見られない特徴を示し、古人類学者はこれらの化石が人類系の初期のメンバーを表していると推測するように導いた。, ヒトの化石記録に現れる最初の人間のような形質は、二足歩行とより小さい鈍い犬歯です。
現在知られている最も古いヒト類は、ChadからのSahelanthropus tchadensisである(Brunet et al. 2005)およびケニアからのOrrorin tugenensis(Senut et al. 2001). サヘラントロプスは、6-7myaの間にあり、大部分が完全な頭蓋骨とその他の断片的な遺跡から知られている。 その脳の大きさ、360ccは、チンパンジーで見られる範囲内であり、頭蓋骨は、男性のゴリラに似た厚さの巨大な眉の尾根を持っています(Brunet et al. 2005)., しかしながら、脊髄が通る頭蓋骨の基部にある穴である大孔の位置と向きは、Sahelanthropusが立って二足歩行し、脊柱が類人猿や他の四足動物のように水平ではなく、現代人のように垂直に保持されていることを示唆している(Zollikofer et al. 2005). Orrorinは部分的な腿骨を含むpostcranial化石から主に知られています。 大腿骨の近位部分は現代人のものと類似しており、この種が二足歩行であることを示唆している(Pickford et al. 2002)., オルロリンの頭蓋骨は回収されておらず、その頭蓋骨の形態と脳の大きさは不明である。 OrrorinとSahelanthropusの両方で、男性の犬の歯は現代人よりも大きく尖っていますが、男性の類人猿の犬の歯に比べて小さくて鈍いです。 このことは、イヌの性的二型性、そしてひいては、雌への交配アクセスのための男性間の競争が、これらの初期のヒト族では、大類人猿と比較して減少していたことを示唆している。
はるかに最もよく知られている初期のヒト人は、Ardipithecus ramidus、a4です。,ほぼ完全な骨格だけでなく、他の多数の歯科および骨格遺跡から知られているエチオピアからの4万年前の種(White et al. 2009). アール ramidusおよび断片的な遺跡から知られている古い、関連する種、Ar。 kadabba(5.8-5.2mya)は、OrrorinとSahelanthropusのものと同様に減少した犬歯を持っています。 Arの頭蓋骨。 ラミドゥスはかなり類人猿に似ており、サヘラントロプスのそれに大きく似ており、300-350ccの小さなチンパンジーサイズの脳を持っています(図2)。 アルディピテクス後頭蓋骨骨格は興味深いです。, ひどく断片化されたが、回収された骨盤は生きている類人猿の形態とは全く異なる形態を明らかにし、Ardipithecusが二足歩行したことを強く示唆するより短く、よりボウルのような形状を示している。 しかし、その長い前肢と指とその発散、最初のつま先(外反母趾)をつかむことは、Ardipithecusが木の中でその時間の多くを費やしたことを示唆しています。 全体的な印象は、それが地面にあえていつでも二足歩行歩いた主に樹上種のものです。,
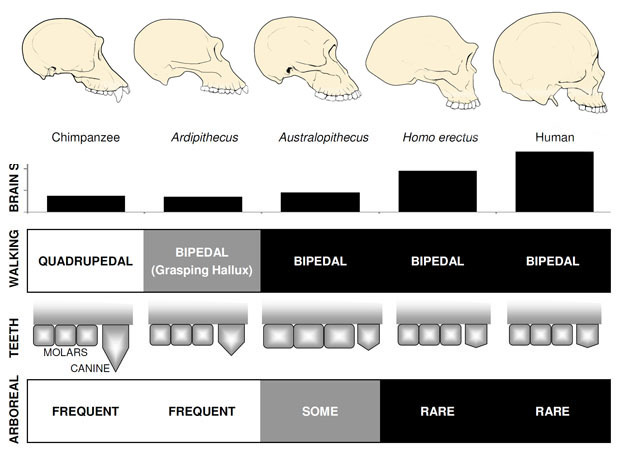
Australopithecus
4mya頃には、陸上二足歩行に熟達していたが、食物と保護のために木を使用し続けたヒトであるAustralopithecus属の最も初期のメンバーが見つかった。 アウストラロピテクスの最初の標本は1924年に南アフリカで発見され(Dart、1925)、その後の八十年にわたる研究努力は、東アフリカと南アフリカのすべてのサイトでいくつかの種から数百の化石を生産している。, 我々は今、アウストラロピテクスはほぼ三百万年間持続する非常に成功した属であったことを知っている(図1)。
最もよく知られているアウストラロピテクス種は、東アフリカからのA.afarensis(3.6-2.9mya)と南アフリカからのA.africanus(3.2–2.0mya)です。, これらの種の骨盤と下肢は、彼らが完全に二足歩行であったことを明確に示しています:骨盤は短く、ボウル型であり、二足歩行中の体幹安定化のために、現代人のように臀部の筋肉を体の側面に持ち込み、最初のつま先は他のつま先と一致しています(Ward、2002;Harcourt-Smith and Aiello、2004)。 アウストラロピテクスの足は、中足骨と化石化されたラエトリの足跡の分析に基づいて、人間のような弓を持っていたかもしれない(Ward et al. 2011)., それにもかかわらず、現代人と比較して、前腕は長く、指とつま先は長く、やや湾曲しており、アウストラロピテクスは定期的に夜間に捕食者からの避難として飼育するために木を使用していたことを示唆している。 この混合地上&樹上戦略は、彼らが住んでいた混合森林とサバンナ環境でよくこれらの種を提供しているだろう。
アウストラロピテクスの脳の大きさは、チンパンジーやゴリラと同様に390-515ccの範囲であった(Falk et al., 2000)、認知能力は生きている類人猿に広く類似していたことを示唆している(図2)。 アウストラロピテクスの体の大きさはかなり小さく、性的に二形であり、女性では約30kg、男性では40kgであった(McHenry、1992)。 このレベルの二型性は、以前のヒト科のように小さく、鈍く、単型であったイヌ科には反映されない。
犬歯とは異なり、アウストラロピテクスの大臼歯は以前のヒト科の歯よりもはるかに大きく、エナメル質が厚かった。 これを食付属、質の低い工場食を必要とした強力な咀嚼プラグインの実行部になります。, 巨大な歯と咀嚼筋のために”堅牢な”アウストラロピテクス(しばしば別の属Paranthropusによって分類される)として知られているアウストラロピテクスのサブグループは、この適応を極端に取った。 ほとんどのアウストラロピテクス種は2myaによって絶滅したが、いくつかの堅牢な形態は東アフリカと南アフリカで約1.2myaまで持続した。
ホモ属
私たち自身の属であるホモの最も初期の化石は、東アフリカで発見され、2.3myaに日付を記入しています(Kimbel et al. 1997)., これらの初期の標本は、脳と体の大きさがアウストラロピテクスと似ていますが、臼歯の違いを示し、食事の変化を示唆しています。 確かに、少なくとも1.8myaによって、私たちの属の初期のメンバーは、原始的な石器を使って動物の死体を肉屋にし、エネルギーが豊富な肉と骨髄を植物ベースの食事に加えていました。
ホモ属の最も古いメンバー、H.habilis(2.3–1.4mya)は東アフリカで発見され、屠殺された動物の骨や単純な石器に関連しています(Blumenschine et al. 2003). そのより手ごわいと広範な子孫、H。, erectusは、アフリカとユーラシア全域で発見され、1.9myaから100kyaまで持続し、おそらくそれ以降でも持続します(Anton、2003)。 H.erectusは、現代人と同様に、アウストラロピテクスに見られる登山のための前肢の適応を欠いていました(図2)。 その世界的な拡大はh.erectusが非常に異なった環境で合わせ、繁栄する認識能力と生態学的に適用範囲が広かったことを提案する。 驚くことではないが、H.erectusでは、後のアジアの標本では1,250ccまでの脳の大きさが大幅に増加し始めている(Anton、2003)。 モルサイズはHで縮小されます。, エレクトゥスはアウストラロピテクスに比べて柔らかく豊かな食事を反映している。
約700kya、そしておそらく以前に、アフリカのH.erectusは、体の割合、歯の適応、および認知能力の点で私たちと非常によく似た種であるh.heidelbergensisを生じさせた(Rightmire、2009)。 H.heidelbergensisは、しばしば”古風な”ホモ-サピエンスと呼ばれ、活発なビッグゲームハンターであり、洗練されたLevalloisスタイルのツールを生産し、少なくとも400kyaが火を制御することを学んだ(Roebroeks and Villa、2011)。 ネアンデルタール人(H., ネアンデルタレンシス)、頑丈な体格、複雑な行動、および私たちのサイズに似た脳を持つ冷たい適応ヒト族は、少なくとも250kya(Rightmire、2008;Hublin、2009)によってヨーロッパのH.heidelbergensis集団から進化したと考えられている。
化石とDNAの証拠は、私たち自身の種であるH.sapiensは、おそらくH.heidelbergensisからアフリカ200kya(Relethford、2008;Rightmire、2009)で進化したことを示唆しています。 H.の増加された行動の洗練。, サピエンスは、私たちの大きな脳(1,400cc)とより広範なツールセットと巧妙な狩猟技術の考古学的証拠によって示されているように、私たちの種はアフリカ大陸で繁栄し成長することを可能にしました。 100kyaまでに、私たちの種はユーラシアに流出し、最終的には世界中でオーストラリアとアメリカ大陸に拡大しました(DiGiorgio et al. 2009). 私たちの種は、ヨーロッパのネアンデルタール人やアジアの同様の形態を含む、彼らが遭遇した他のヒト族を置き換えた道に沿って。 (すべてがデータのこの解釈に同意するわけではないことに注意してください。TryonとBaileyを参照)。, ネアンデルタール人の化石から抽出された古代のDNAの研究は、我々の種が時折彼らと交配した可能性があることを示唆している(Green et al., 2010). 農業や都市化などの文化的革新が私たちの周りの風景や種を形作るように、私たちの増加する世界的な影響は今日も続いています。
概要
類人猿のような中新世の祖先からの私たちの種の進化は複雑なプロセスでした。 私たちの系統は側枝と進化の行き止まりでいっぱいであり、数百万年以上持続していた堅牢なアウストラロピスのような種は消え去っています。, 二足歩行のようないくつかの人間の特徴は非常に早く進化しましたが、大きな脳のような他のものは比較的最近まで進化しませんでした。 モルサイズのようなさらに他の形質は、生態学的圧力を変えることによって後で押し戻されるために一方向に進化した。 いくつかの事前に決定された目的地に向かってまっすぐなコースをチャート化する強力な船ではなく、私たちの系統の進化-確かに、あらゆる種の系統の—環境変化、遺伝的運、および地質学的なチャンスの変化する海によって投げられた救命ボートのイメージに合っています。 一つは、次の六百万年が私たちを取るかもしれない場所を不思議。