Introduzione
La grande intuizione di Darwin, e il principio unificante della biologia oggi, è che tutte le specie sono legate l’una all’altra come sorelle, cugini e parenti lontani in un vasto albero genealogico della vita. Le implicazioni sono mozzafiato; se potessimo viaggiare indietro abbastanza nel tempo, troveremmo antenati comuni tra noi e ogni altro organismo vivente, dagli istrici ai fenicotteri ai cactus., La nostra famiglia evolutiva immediata è composta dagli ominoidi, il gruppo di primati che comprende le ” scimmie minori “(siamang e gibboni) e le” grandi scimmie ” (scimpanzé, bonobo, gorilla e oranghi). Tra le grandi scimmie, i nostri parenti più stretti sono gli scimpanzé e i bonobo (Figura 1). I reperti fossili, insieme a studi sul DNA umano e delle scimmie, indicano che gli umani condividevano un antenato comune con scimpanzé e bonobo circa 6 milioni di anni fa (mya)., Iniziamo questa discussione sull’evoluzione della nostra specie in Africa, verso la fine del periodo geologico noto come Miocene, poco prima che il nostro lignaggio divergesse da quello degli scimpanzé e dei bonobo.
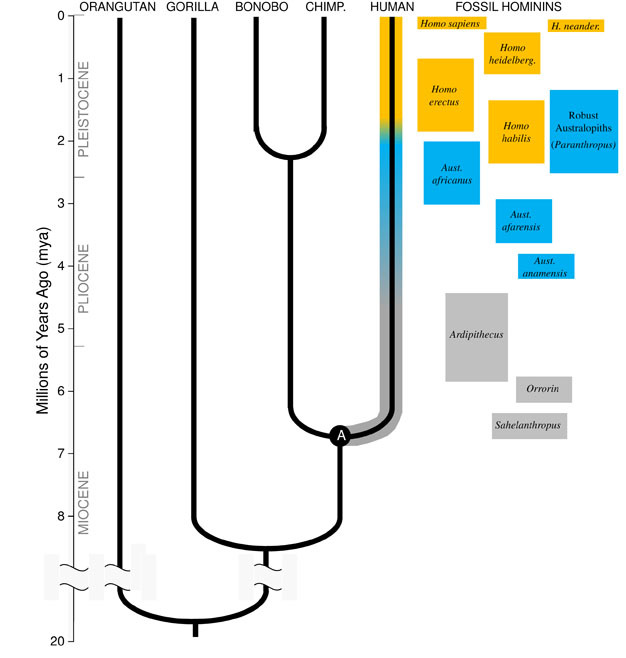
Origini mioceniche del Lignaggio degli Ominidi
Per comprendere l’evoluzione di qualsiasi specie, dobbiamo prima stabilire il suo stato ancestrale: da che tipo di animale si è evoluto? Per il nostro lignaggio, questo richiede che cerchiamo di ricostruire l’Ultimo antenato comune di umani e scimpanzé (contrassegnato “A” in Figura 1). L’Uomo-Scimpanzé Ultimo antenato comune (HC-LCA) è la specie da cui il lignaggio degli ominidi e lo scimpanzé & lignaggio bonobo divergevano., Gli ominidi sono specie sul nostro ramo dell’albero ominoide dopo la divisione con lo scimpanzé & linea bonobo, incluse tutte le specie estinte e i rami laterali evolutivi (Figura 1).
C’era una grande diversità di specie di scimmie nel Miocene, con dozzine di specie conosciute dai reperti fossili in Africa, Europa e Asia. Queste specie variavano nella loro anatomia ed ecologia, e non è chiaro quale, se del caso, delle specie fossili scoperte finora rappresentino l’HC-LCA (Kunimatsu et al. 2007; Young e MacLatchy, 2004)., Tuttavia, sappiamo da prove fossili e comparative che era molto più simile alle scimmie viventi che agli esseri umani viventi. L’HC-LCA avrebbe avuto un cervello e un corpo di dimensioni scimmiesche, con braccia e dita relativamente lunghe e un piede che gli permetteva di foraggiare tra gli alberi. I denti canini erano probabilmente grandi e affilati, come visto in diversi ominoidi del Miocene. Inoltre, i canini erano probabilmente sessualmente dimorfici, con i maschi che avevano canini molto più grandi delle femmine, come visto tra le grandi scimmie viventi e fossili del Miocene., Come le scimmie viventi avrebbe camminato quadrupedi (a quattro zampe) quando era a terra, e la sua dieta sarebbe consistita quasi interamente di alimenti vegetali, principalmente frutta e foglie.
I primi ominidi
I cambiamenti da un’anatomia simile a una scimmia sono distinguibili nei fossili di ominidi del tardo Miocene in Africa. Alcune specie ominoidi di questo periodo mostrano tratti tipici degli umani ma non si vedono nelle altre scimmie viventi, portando i paleoantropologi a dedurre che questi fossili rappresentano i primi membri del lignaggio degli ominidi., I primi tratti umani ad apparire nei reperti fossili di ominidi sono la camminata bipede e i canini più piccoli e smussati.
I più antichi ominidi attualmente conosciuti sono Sahelanthropus tchadensis del Ciad (Brunet et al. 2005) e Orrorin tugenensis dal Kenya (Senut et al. 2001). Sahelanthropus, datato tra 6 e 7 mya, è noto da un cranio in gran parte completo e alcuni altri resti frammentari. La sua dimensione del cervello, 360cc, è all’interno della gamma visto in scimpanzé, e il cranio ha una cresta fronte massiccia, simile nello spessore di gorilla maschi (Brunet et al. 2005)., Tuttavia, la posizione e l’orientamento del forame magnum, il foro nella base del cranio attraverso il quale passa il midollo spinale, suggerisce che Sahelanthropus stava e camminava bipedi, con la sua colonna vertebrale tenuta verticalmente come negli esseri umani moderni piuttosto che orizzontalmente come nelle scimmie e altri quadrupedi (Zollikofer et al. 2005). Orrorin è conosciuto principalmente da fossili postcranici, tra cui un femore parziale. La porzione prossimale del femore mostra somiglianze con quelle degli esseri umani moderni, suggerendo che la specie era bipede (Pickford et al. 2002)., Nessun teschio di Orrorin è stato recuperato, e quindi la sua morfologia cranica e le dimensioni del cervello sono incerte. In entrambi Orrorin e Sahelanthropus i denti canini dei maschi sono più grandi e più appuntiti che negli esseri umani moderni, ma sono piccoli e smussati rispetto ai canini delle scimmie maschi. Ciò suggerisce che il dimorfismo sessuale canino — e per estensione, la competizione tra i maschi per l’accesso all’accoppiamento alle femmine-era diminuita in questi primi ominidi rispetto alle grandi scimmie.
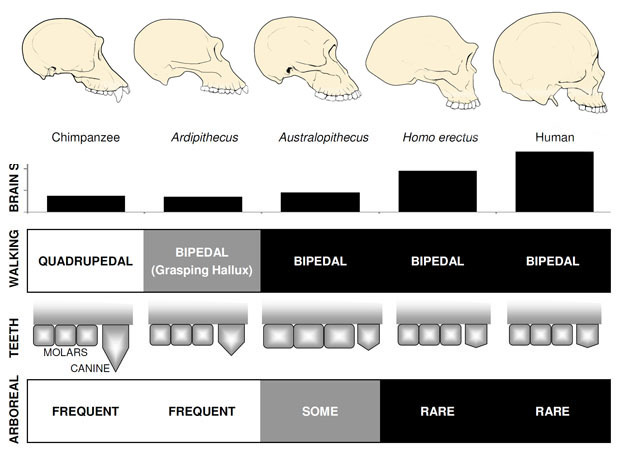
Di gran lunga il più noto ominide precoce è Ardipithecus ramidus, a 4.,specie di 4 milioni di anni dall’Etiopia, che è conosciuta da uno scheletro quasi completo e da numerosi altri resti dentali e scheletrici (White et al. 2009). AR. ramidus e un vecchio, specie correlate noti da resti frammentari, Ar. i kadabba (5,8–5,2 mya), hanno canini ridotti simili a quelli di Orrorin e Sahelanthropus. Il cranio di Ar. ramidus è piuttosto simile a una scimmia e sostanzialmente simile a quello di Sahelanthropus, con un cervello di piccole dimensioni di scimpanzé di 300-350cc (Figura 2). Lo scheletro postcranico di Ardipithecus è intrigante., Anche se gravemente frammentato, il bacino recuperato rivela una morfologia molto diversa da quella delle scimmie viventi, con una forma più corta, più a forma di ciotola che suggerisce fortemente Ardipithecus camminato bipede; questo è coerente con la posizione del forame magnum, che suggerisce una postura eretta. Tuttavia, i suoi lunghi arti anteriori e le dita e il suo primo alluce divergente e afferrante suggeriscono che Ardipithecus trascorresse gran parte del suo tempo tra gli alberi. L’impressione generale è di una specie in gran parte arboricola che camminava bipede ogni volta che si avventurava a terra.,
Australopithecus
Intorno a 4mya troviamo i primi membri del genere Australopithecus, ominidi che erano abili bipedi terrestri ma continuavano ad usare gli alberi per cibo e protezione. I primi esemplari di Australopithecus furono scoperti in Sud Africa nel 1924 (Dart, 1925), e gli sforzi di ricerca nei successivi otto decenni hanno prodotto centinaia di fossili da diverse specie in siti in tutta l’Africa orientale e meridionale., Ora sappiamo che Australopithecus era un genere di grande successo che persistette per quasi tre milioni di anni (Figura 1).
Le specie Australopithecus più note sono A. afarensis (3,6-2,9 mya) dell’Africa orientale e A. africanus (3,2–2,0 mya) del Sudafrica., Il bacino e l’arto inferiore di queste specie indicano chiaramente che erano completamente bipedi: il bacino è corto e a forma di ciotola, portando i muscoli glutei intorno al lato del corpo, come negli esseri umani moderni, per la stabilizzazione del tronco durante il bipedismo, e il primo dito è in linea con le altre dita (Ward, 2002; Harcourt-Smith e Aiello, 2004). Il piede dell’Australopithecus potrebbe anche avere avuto un arco simile a quello umano, sulla base dell’analisi dei metatarsi e delle impronte fossili di Laetoli (Ward et al. 2011)., Tuttavia, rispetto agli umani moderni, gli avambracci erano lunghi e le dita delle mani e dei piedi erano lunghi e un po ‘ ricurvi, suggerendo che l’Australopithecus usasse regolarmente gli alberi per foraggiare e forse come rifugio dai predatori durante la notte. Questa strategia arborea terrestre mista & avrebbe servito bene queste specie negli ambienti boschivi misti e nella savana in cui abitavano.
La dimensione del cervello in Australopithecus variava tra 390 e 515cc, simile a scimpanzé e gorilla (Falk et al., 2000), suggerendo che le capacità cognitive erano sostanzialmente simili alle scimmie viventi (Figura 2). Le dimensioni del corpo nell’Australopithecus erano piuttosto piccole e sessualmente dimorfiche, circa 30 kg per le femmine e 40 kg per i maschi (McHenry, 1992). Questo livello di dimorfismo non si riflette nei canini, che erano piccoli, smussati e monomorfi come nelle precedenti ominine.
A differenza dei canini, i denti molari nell’Australopithecus erano molto più grandi di quelli degli ominidi precedenti e avevano uno smalto più spesso. Ciò suggerisce che la loro dieta includeva cibi vegetali duri e di bassa qualità che richiedevano una masticazione potente da elaborare., Un sottogruppo di Australopithecus, noto come “robusto” australopiths (spesso etichettato da un genere separato Paranthropus) a causa dei loro enormi denti e muscoli masticatori, ha preso questo adattamento all’estremo. La maggior parte delle specie di Australopithecus erano estinte da 2 mya, ma alcune forme robuste persistevano fino a circa 1,2 mya in Africa orientale e meridionale.
Il Genere Homo
I primi fossili del nostro genere, Homo, si trovano in Africa orientale e datati a 2,3 mya (Kimbel et al. 1997)., Questi primi esemplari sono simili nelle dimensioni del cervello e del corpo a Australopithecus, ma mostrano differenze nei loro denti molari, suggerendo un cambiamento nella dieta. Infatti, di almeno 1,8 mya, i primi membri del nostro genere usavano strumenti di pietra primitivi per macellare carcasse di animali, aggiungendo carne e midollo osseo ricchi di energia alla loro dieta a base vegetale.
Il più antico membro del genere Homo, H. habilis (2.3–1.4 mya) si trova in Africa orientale ed è associato a ossa di animali macellati e semplici strumenti di pietra (Blumenschine et al. 2003). Il suo discendente più formidabile e diffuso, H., erectus, si trova in tutta l’Africa e l’Eurasia e persisteva da 1,9 mya a 100 kya, e forse anche più tardi (Anton, 2003). Come gli esseri umani moderni, H. erectus mancava gli adattamenti anteriori per l’arrampicata visto in Australopithecus (Figura 2). La sua espansione globale suggerisce H. erectus era ecologicamente flessibile, con la capacità cognitiva di adattarsi e prosperare in ambienti molto diversi. Non sorprendentemente, è con H. erectus che iniziamo a vedere un importante aumento delle dimensioni del cervello, fino a 1.250 cc per gli esemplari asiatici successivi (Anton, 2003). La dimensione molare è ridotta in H., erectus rispetto all’Australopithecus, che riflette la sua dieta più morbida e ricca.
Circa 700 kya, e forse prima, H. erectus in Africa ha dato origine a H. heidelbergensis, una specie molto simile a noi in termini di proporzioni corporee, adattamenti dentali e capacità cognitive (Rightmire, 2009). H. heidelbergensis, spesso definito un Homo sapiens “arcaico”, era un cacciatore di grandi dimensioni attivo, produceva sofisticati strumenti in stile Levallois e da almeno 400 kya aveva imparato a controllare il fuoco (Roebroeks e Villa, 2011). Neanderthal (H., neanderthalensis), ominidi adattati al freddo con fisici robusti, comportamenti complessi e cervelli di dimensioni simili al nostro, si pensa che si siano evoluti dalle popolazioni di H. heidelbergensis in Europa di almeno 250 kya (Rightmire, 2008; Hublin, 2009).
Le prove fossili e del DNA suggeriscono che la nostra specie, H. sapiens, si è evoluta in Africa 200 kya (Relethford, 2008; Rightmire, 2009), probabilmente da H. heidelbergensis. L’accresciuta sofisticazione comportamentale di H., sapiens, come indicato dai nostri grandi cervelli (1.400 cc) e prove archeologiche di un set di strumenti più ampio e tecniche di caccia intelligenti, hanno permesso alla nostra specie di prosperare e crescere nel continente africano. Con 100kya, la nostra specie si riversò in Eurasia, espandendosi infine in tutto il mondo in Australia e nelle Americhe (DiGiorgio et al. 2009). Lungo la strada la nostra specie ha spostato altri ominidi che hanno incontrato, compresi i Neanderthal in Europa e forme simili in Asia. (Si noti che non tutti sono d’accordo con questa interpretazione dei dati, vedere Tryon e Bailey)., Studi di DNA antico estratto da fossili di Neanderthal suggeriscono che la nostra specie potrebbe aver occasionalmente incrociato con loro (Green et al., 2010). Il nostro crescente impatto globale continua ancora oggi, poiché innovazioni culturali come l’agricoltura e l’urbanizzazione modellano il paesaggio e le specie che ci circondano.
Sommario
L’evoluzione della nostra specie da un antenato del Miocene simile a una scimmia è stato un processo complesso. Il nostro lignaggio è pieno di rami laterali e vicoli ciechi evolutivi, con specie come i robusti australopiths che persistettero per oltre un milione di anni prima di svanire., Alcuni tratti umani, come il bipedismo, si sono evoluti molto presto, mentre altri, come i grandi cervelli, non si sono evoluti fino a tempi relativamente recenti. Ancora altri tratti, come la dimensione dei molari, si sono evoluti in una direzione solo per essere respinti più tardi cambiando le pressioni ecologiche. Piuttosto che una potente nave che traccia una rotta dritta verso una destinazione predeterminata, l’evoluzione del nostro lignaggio-anzi, del lignaggio di qualsiasi specie — si adatta all’immagine di una scialuppa di salvataggio lanciata dai mari mutevoli del cambiamento ambientale, della fortuna genetica e del caso geologico. Ci si chiede dove potrebbero portarci i prossimi sei milioni di anni.