Johdanto
Darwinin suuri oivallus, ja yhdistävä periaate biologian tänään, on se, että kaikki lajit ovat sukua toisilleen kuin sisarukset, serkut ja kaukaisten kin laaja perhe-elämän puu. Vaikutukset ovat henkeäsalpaavat; jos voisimme matkustaa takaisin kauas tarpeeksi ajoissa, emme olisi löytää yhteinen esi keskenämme ja jokainen elävä organismi, mistä porcupines että flamingot, jotta kaktukset., Meidän välitöntä evoluution perhe koostuu hominoids, ryhmä kädellisten, joka sisältää ”vähemmän apinoita” (siamangs ja gibbons) sekä ”suuret apinat” (simpanssit, bonobos, gorillat ja orangit). Suurapinoiden joukossa lähimpiä sukulaisiamme ovat simpanssit ja bonobot (Kuva 1). Fossiilisten ennätys, sekä tutkimuksia ihmisen ja apinan DNA: ta, osoittavat, että ihmisillä on ollut yhteinen esi-isä simpanssien kanssa ja bonobos joskus noin 6 miljoonaa vuotta sitten (mya)., Meidän on aloitettava tämä keskustelu lajimme evoluutio Afrikassa, lähellä loppua geologinen ajanjakso tunnetaan Mioseenikautena, juuri ennen meidän linjaa poikkesivat että simpanssien ja bonobos.
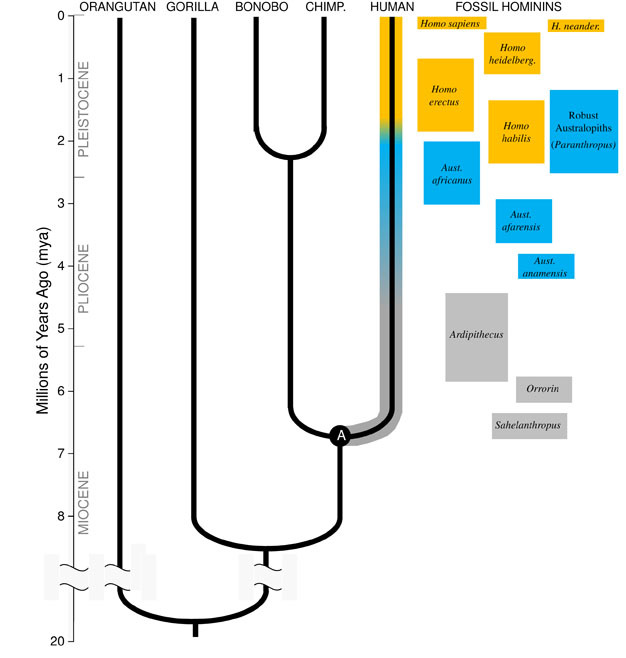
Mioseenikautena Alkuperä Hominin Lineage
jotta ymmärtää kehitystä, lajista, meidän on ensin luotava sen esi-isien tila: millainen eläin teki sen kehittyä? Meidän linjaa, tämä edellyttää, että yritämme rekonstruoida Viimeinen Yhteinen Esi ihmisten ja simpanssien (merkitty ”A” Kuvassa 1). Ihmisen ja Simpanssin Viimeinen Yhteinen Esi-isä (HC-LCA) on laji, josta hominin linjaa ja simpanssi & bonobo linjaa poikkesivat toisistaan., Hominins ovat lajeja, meidän haara hominoid puu splitin jälkeen, jossa simpanssi & bonobo linja, mukaan lukien kaikki sukupuuttoon kuolleita lajeja ja evoluution puolella oksat (Kuva 1).
Siellä oli suuri monimuotoisuus apina lajeja Mioseenikautena, kymmeniä lajeja tunnetaan fossiilisten ennätys kaikkialla Afrikassa, Euroopassa ja Aasiassa. Näiden lajien vaihdellut niiden anatomia ja ekologia, ja se ei ole selvää, mikä, jos mikään, fossiilisia lajeja löydettiin toistaiseksi edustamaan HC-LCA (Kunimatsu et al. 2007; Young and MacLatchy, 2004)., Tiedämme kuitenkin fossiilisten ja vertailevien todisteiden perusteella, että se muistutti paljon enemmän eläviä apinoita kuin eläviä ihmisiä. HC-LCA olisi ollut apina-kokoinen aivojen ja ruumiin, suhteellisen pitkät kädet ja sormet ja ahne, että jalka saa sitä rehua puissa. Koiran hampaat olivat todennäköisesti suuret ja terävät, kuten useissa Mioseenihominoideissa on nähty. Lisäksi koirat olivat luultavasti yliseksuaalisuus, miehillä, joilla on paljon suurempi kulmahampaat kuin naisilla, kuten nähdään joukossa elävien ihmisapinoiden ja Mioseenikautena fossiileja., Kuten elävät apinat se olisi voinut quadrupedally (kontallaan), kun maahan, ja sen ruokavalio olisi koostui lähes kokonaan kasvisruokaa, pääasiassa hedelmiä ja lehtiä.
Alussa Hominins
Muutokset apinamaisen anatomia on havaittavissa hominoid fossiileja myöhään Mioseenikautena Afrikassa. Jotkut hominoid lajin tämän kauden näyttely piirteitä, jotka ovat tyypillisiä ihmisille, mutta ei nähnyt muita eläviä apinoita, johtava paleoanthropologists päätellä, että nämä fossiilit edustavat varhaisia jäseniä hominin linjaa., Ensimmäiset hominiinin fossiilistossa esiintyvät ihmismäiset piirteet ovat kaksijalkainen kävely ja pienemmät, tylpät kulmahampaat.
vanhimmat nykyisin tunnetut hominiinit ovat Tšadista kotoisin oleva Sahelanthropus tchadensis (Brunet et al. 2005) ja Orrorin tugenensis Keniasta (Senut et al. 2001). Sahelanthropus, ajoitettu välillä 6 ja 7 mya, tunnetaan suurelta osin täydellisestä pääkallosta ja joistakin muista fragmentaarisista jäänteistä. Sen aivojen koko, 360cc, on alueella nähty simpanssit, ja kallo on massiivinen otsa ridge, samanlainen paksuus uros gorillat (Brunet ym. 2005)., Kuitenkin, sijainti ja suunta foramen magnum, reikä kallon pohja, jonka kautta selkäydin kulkee, viittaa siihen, että Sahelanthropus seisoi ja käveli bipedally, sen selkärangan pystyasennossa kuin modernin ihmisen sijaan horisontaalisesti kuin apinat ja muut nelijalkaiset (Zollikofer et al. 2005). Orrorin tunnetaan ennen kaikkea kallonjälkeisistä fossiileista, muun muassa osittaisesta reisiluusta. Reisiluun proksimaalisessa osassa on yhtäläisyyksiä nykyihmisten kanssa, mikä viittaa siihen, että laji oli kaksijalkainen (Pickford et al. 2002)., Orroriinin kalloja ei ole löydetty, joten sen kallon morfologia ja aivojen koko ovat epävarmoja. Molemmissa Orrorin ja Sahelanthropus koiran hampaat urokset ovat suurempia ja enemmän terävä kuin nykyajan ihmiset, mutta on pieni ja tylsä verrattuna kulmahampaat mies apinoita. Tämä viittaa siihen, että koiran seksuaalinen dimorphism — ja laajennus, kilpailu miesten keskuudessa pariutumisen pääsy naisilla — oli vähentynyt nämä varhaiset hominins verrattuna ihmisapinoita.
ylivoimaisesti tunnetuin varhainen hominiini on Ardipithecus ramidus, a 4.,4 miljoonaa vuotta vanhaa lajia Etiopiasta, joka tunnetaan lähes täydellisestä luurangosta sekä lukuisista muista hammas – ja luurankojäännöksistä (White et al. 2009). A. ramidus ja vanhempi, sukua oleva laji tunnetaan fragmentaarisista jäänteistä, Ar. kadabballa (5,8-5,2 mya) on orrorinin ja Sahelanthropuksen kaltaisia pienennettyjä kulmahampaita. Arin kallo. ramidus on melko apinan ja pitkälti samanlainen kuin Sahelanthropus, pieni simpanssi-kokoinen aivot 300–350cc (Kuva 2). Ardipithecus postcranial luuranko on kiehtova., Vaikka pahasti hajanainen, lantion talteen paljastaa morfologia on varsin erilainen kuin elävät apinat, lyhyempi, enemmän kulho-muoto, joka viittaa vahvasti siihen, Ardipithecus käveli bipedally; tämä on sopusoinnussa foramen magnum-asema, joka viittaa siihen, pystyasennossa. Kuitenkin, sen pitkän forelimbs ja sormien ja sen erilaiset, ahne ensimmäinen kärki (hallux) viittaavat siihen, Ardipithecus vietti paljon aikaa puissa. Yleisvaikutelma on paljolti arboreaalisesta lajista, joka käveli kaksijakoisesti aina, kun se uskaltautui maahan.,
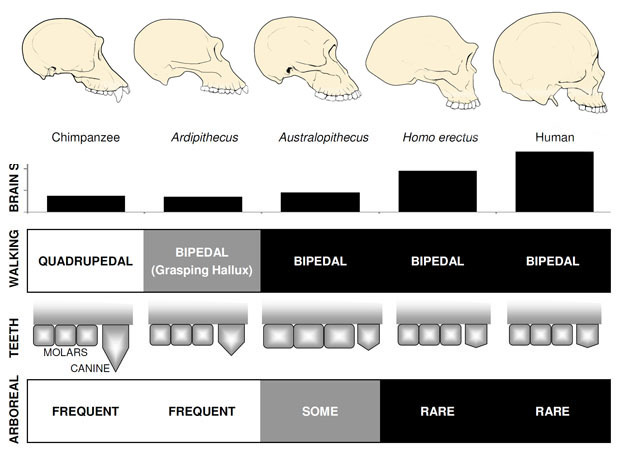
Australopithecus
Noin 4mya löydämme aikaisintaan jäsenet suvun Australopithecus, hominins, jotka olivat taitavia maanpäällisen bipeds, mutta edelleen käyttää puita, ruokaa ja suojaa. Ensimmäinen yksilöt Australopithecus löydettiin Etelä-Afrikasta vuonna 1924 (Tikka, 1925), ja tutkimusta seuraavien kahdeksan vuosikymmentä on tuottanut satoja fossiileja useita lajeja sivustoja ympäri Itä-ja Etelä-Afrikassa., Tiedämme nyt, että Australopithecus oli erittäin onnistunut suvun, joka kesti lähes kolme miljoonaa vuotta (Kuva 1).
tunnetuin Australopithecus-lajit ovat A. afarensis (3.6–2.9 mya) Itä-Afrikan ja A. africanus (3.2–2.0 mya) Etelä-Afrikasta., Lantion ja alaraajojen näistä lajeista selvästi osoittaa, että he olivat täysin bipedal: lantio on lyhyt ja kulho-muotoinen, jolloin pakaroiden lihaksia noin puolella kehon, kuten nykyajan ihmiset, runko vakauttaminen aikana bipedalism, ja ensimmäinen kärki on linjassa muiden varpaat (Ward, 2002; Harcourt-Smith ja Aiello, 2004). Että Australopithecus jalka voi jopa on ollut ihmisen kaltainen kaari, joka perustuu analyysi metatarsals ja kivettyneet Laetoli jalanjäljet (Ward et al. 2011)., Kuitenkin, verrattuna nykyajan ihmiset, käsivarret olivat pitkiä ja sormet ja varpaat olivat pitkä ja hieman kaareva, mikä viittaa siihen, että Australopithecus käytetään säännöllisesti puiden rehu ja ehkä turvaa pedoilta yöllä. Tämä sekoittaa maanpäällisen & puissa strategia olisi toiminut näiden lajien hyvin sekoitettu metsää ja savannah ympäristöissä, ne asuttu.
Aivojen koko Australopithecus vaihteli välillä 390 ja 515cc, samanlainen kuin simpanssit ja gorillat (Falk et al., 2000), mikä viittaa kognitiivisiin kykyihin olivat suurin piirtein samanlaisia kuin elävät apinat (kuva 2). Kehon koko Australopithecus oli melko pieni ja yliseksuaalisuus, noin 30kg naisilla ja 40kg miehillä (McHenry, 1992). Tämän tason dimorphism ei näy kulmahampaat, jotka olivat pieniä, tylppä, ja monomorfinen kuten aiemmin hominins.
toisin Kuin kulmahampaat, poskihampaat vuonna Australopithecus oli paljon suurempi kuin aikaisemmin hominins, ja oli paksumpi emali. Tämä viittaa siihen, että heidän ruokavalionsa sisälsi kovia, huonolaatuisia kasviruokia, jotka vaativat tehokasta pureskelua prosessoidakseen., Alaryhmä Australopithecus, joka tunnetaan nimellä ”vahva” australopiths (usein merkitty erillisellä suvun Paranthropus), koska niiden valtava hampaat ja poskilihakset, otti tämä sopeutuminen äärimmilleen. Useimmat Australopithecus-lajit kuolivat sukupuuttoon 2 mya, mutta selkeitä muotoja jatkui kunnes noin 1,2 mya Itä-ja Etelä-Afrikassa.
Suvun, Homo
aikaisintaan fossiileja oman suvun, Homo, löytyy Itä-Afrikassa ja päivätty 2.3 mya (Kimbel et al. 1997)., Nämä varhaiset yksilöt muistuttavat aivo-ja ruumiinkooltaan Australopithecusta, mutta niissä on eroja poskihampaissa, mikä viittaa ruokavalion muutokseen. Todellakin, ainakin 1,8 mya, varhainen jäsenet suvun käyttivät alkukantainen pelinappulaa työkalut teurastaja eläinten ruhot, lisäämällä energia-rikas lihaa ja luuytimen niiden kasvi-pohjainen ruokavalio.
vanhin sukuun Homo, H. habilis (2.3–1.4 mya) löytyy Itä-Afrikassa ja liittyy teurasti eläinten luita ja yksinkertainen kivi työkaluja (Blumenschine et al. 2003). Sen pelottavampi ja laajalle levinnyt jälkeläinen H., erectus, löytyy kaikkialla Afrikassa ja Euraasiassa ja jatkui 1,9 mya 100 kya, ja ehkä jopa myöhemmin (Anton, 2003). Kuten nykyajan ihmiset, H. erectus puuttui eturaajan järjestelyt kiipeilyä nähty Australopithecus (Kuva 2). Sen maailmanlaajuinen laajeneminen viittaa siihen, H. erectus oli ekologisesti joustava kognitiivinen kyky sopeutua ja menestyä huomattavasti eri ympäristöissä. Ei ole yllättävää, se on H. erectus, että me alamme nähdä merkittävä kasvattaa aivojen kokoa, jopa 1250 cc myöhemmin Aasian yksilöt (Anton, 2003). Moolikoko pienenee H: ssa., erectus suhteessa Australopithecus, heijastaa sen pehmeämpi, rikkaampi ruokavalio.
Noin 700 kya, ja ehkä aikaisemmin, H. erectus Afrikassa synnytti H. heidelbergensis, laji hyvin paljon meidän kannalta kehon mittasuhteet, hampaiden mukautukset, ja kognitiivinen kyky (Rightmire, 2009). H. heidelbergensis, usein kutsutaan kuin ”arkaainen” Homo sapiens, oli aktiivinen iso peli metsästäjä, valmistettu hienostunut Levallois tyyli työkaluja, ja vähintään 400 kya oli oppinut hallitsemaan tulta (Roebroeks ja Villa, 2011). Neandertalilaiset (H., neanderthalensis), kylmä-mukautettu hominins stout physiques, monimutkaisia toimintoja, ja aivot vastaavat kooltaan meidän, uskotaan ovat kehittyneet H. heidelbergensis populaatiot Euroopassa vähintään 250 kya (Rightmire, 2008; Hublin, 2009).
Fossiilisten ja DNA-todisteet viittaavat siihen, omat lajit, H. sapiens, kehittyi Afrikassa 200 kya (Relethford, 2008; Rightmire, 2009), luultavasti H. heidelbergensis. H., sapiens, kuten meidän suuret aivot (1,400 cc) ja arkeologisia todisteita laajemman työkalu asettaa ja taitava metsästys tekniikoita, saa meidän laji kukoistaa ja kasvaa Afrikan mantereella. By 100kya, lajimme valunut Euraasiassa, lopulta laajentaa koko maailmaa Australiassa ja Amerikassa (DiGiorgio et al. 2009). Matkan varrella lajimme syrjäytti muut kohtaamansa hominiinit, kuten Neandertalilaiset Euroopassa ja vastaavat muodot Aasiassa. (Huomaa, että kaikki eivät ole samaa mieltä tästä tietojen tulkinnasta, KS. Tryon ja Bailey)., Neandertalin fossiileista saadun muinaisen DNA: n tutkimukset viittaavat siihen, että lajimme on saattanut joskus risteytyä niiden kanssa (Green et al., 2010). Lisääntyvä globaali vaikutuksemme jatkuu nykyäänkin, kun Kulttuuriset innovaatiot, kuten maatalous ja kaupungistuminen muokkaavat maisemaa ja lajeja ympärillämme.
Tiivistelmä
evoluutiota lajin apina-kuten Mioseenikautena esi-isä oli monimutkainen prosessi. Meidän suku on täynnä puolella oksat ja evoluution umpikujia, jossa lajit kuten vankka australopiths, jotka jatkuivat yli miljoona vuotta ennen hiipumassa., Jotkut ihmisen piirteet, kuten bipedalismi, kehittyivät hyvin varhain, kun taas toiset, kuten suuret aivot, kehittyivät vasta suhteellisen äskettäin. Vielä muita piirteitä, kuten poskihammas, kehittynyt vain yhteen suuntaan työntää takaisin myöhemmin muuttamalla ekologisia paineita. Sen sijaan voimakas alus kartoitus suora kurssi kohti joitakin ennalta määritetty kohde, evoluutiota linjaa — itse asiassa minkä tahansa lajin lineage — sopii kuvan pelastusvene heitelleet siirtää meret ympäristön muutos, geneettisen onnea, ja geologinen mahdollisuus. Mietitään, mihin seuraavat kuusi miljoonaa vuotta vievät.