Kuva 4.1 (ks. suurennettu)
näin Ollen ymmärrystä tämän synapse johtaa ymmärrystä muille. Siksi keskustelemme ensin synaptisen transmission prosessista luuston neuromuskulaarisessa yhtymäkohdassa.,
neuromuskulaarisen liittymän synaptisen liittymän piirteet näkyvät kuvassa vasemmalla. Luuston lihassyiden hermottamiin moottori neuronien, joiden solu elimet sijaitsevat vatsanpuoleinen sarvi selkäydintä. Aksonin terminaalinen alue synnyttää hyvin hienoja prosesseja, jotka kulkevat luurankolihassoluja pitkin. Näiden prosessien varrella on erikoistuneita rakenteita, joita kutsutaan synapseiksi. Erityisesti synapsien välillä spinal motor neuron ja luuston lihasten solu on nimeltään motor endplate koska sen erityinen rakenne.,
synapse hermo-lihasliitoksessa on kolme tunnusomaiset piirteet kemiallisten synapsien hermoston. Ensinnäkin presynaptisen ja postsynaptisen kalvon välillä on selvä ero. Kaksikon välinen tila tunnetaan synaptisena klefttinä. Tila kertoo meille, siellä on joitakin välittäjä signalointi mekanismi välillä presynaptinen neuroni ja postsynaptisten hermopäätteiden jotta tiedonkulku eri synapsiraossa. Toiseksi on ominaista suuri tiheys pieniä pallomaisia rakkuloita., Nämä synaptiset rakkulat sisältävät välittäjäaineita. Synapseihin liittyy myös suuri mitokondrioiden tiheys. Kolmanneksi, useimmissa tapauksissa, on ominaista paksuuntuminen postsynaptisiin kalvo, joka johtuu ainakin osittain siitä, että postsynaptisiin kalvo on korkea tiheys erikoistuneita reseptoreita, jotka sitovat kemialliset lähetin aineita vapautuu presynaptisen hermosolun. Lisätietoja morfologiset ominaisuudet synaptic liittymissä on Luvussa 8 ja Luvussa 10.
4.,2 Fysiologia Synaptic Siirto Hermo-lihasliitoksessa
Kuva 4.2
kuva oikealla havainnollistaa hyvin kaavamainen tapa, miten se on mahdollista tutkimus fysiologian synaptic siirto luuston lihasliitos yksityiskohtaisesti. Pala lihasta ja siihen kiinnittynyt hermo sijoitetaan pieneen koekammioon, joka on täytetty sopivalla Ringer-liuoksella. Lihassolun lepopotentiaali kirjataan mikroelektrodilla. Elektrodit asetetaan myös hermoaksonin pinnalle., Lyhyet sähköiskut aiheuttavat aktivointimahdollisuuksia, jotka etenevät synaptiseen terminaaliin.
alla Oleva kuva havainnollistaa kahden tyyppisiä mahdollisia muutoksia, jotka kirjataan tällainen yksittäinen hermo-lihas valmistelu. Kokeilu osoittaa myös ominaisuuksia voimakas lääke, kurare, joka on osoittautunut erittäin hyödyllinen opiskelu prosessi synaptic siirto luuston lihasliitos. A-osa havainnollistaa sitä, missä järjestyksessä mahdolliset muutokset kirjataan lihassoluun motorisen aksonin stimuloinnin seurauksena., Nuoli osoittaa ajankohdan, jolloin isku toimitetaan moottoriaksonille. Huomaa, että shokin jälkeen on hiljaista aikaa. Viive johtuu siitä, että aika kuluu aktiopotentiaalin moottorin axon levittää sen päällä aloittamista. Viiveen jälkeen lihassoluun kirjataan kahdenlaisia potentiaaleja. Ensinnäkin seuraavassa keskustelussa keskitytään suhteellisen hitaasti muuttuviin mahdollisuuksiin., Jos on hidas, alkuperäisen potentiaali on riittävän suuri, koska se yleensä on luuston lihaksen soluissa, toinen mahdollinen, toiminta mahdollisuudet on herättänyt lihassolujen.
Kuva 4.3
aktiopotentiaalien luuston lihaksen soluissa ovat, koska ionic mekanismit vastaavat keskusteltu aiemmin. Erityisesti na+-läpäisevyydessä tapahtuu jännitteestä riippuva muutos, jota seuraa k+ – läpäisevyyden viivästynyt kasvu. (Sileiden lihassolujen ja sydänlihassolujen ionimekanismit ovat kuitenkin erilaiset.,)
taustalla Oleva tapahtuma, joka laukaisee aktiopotentiaalin voidaan paljastaa ottamalla etu kuraren nuoli myrkkyä käytetään joissakin Etelä-Amerikan Intiaanit. Pieni annos curarea (osa B) vähentää taustalla olevaa tapahtumaa, mutta sitä ei silti alenneta riittävästi, jotta se alittaisi kynnyksen. Jos hieman suurempi annos kuraren on toimitettu (Osa C), hidas taustalla oleva tapahtuma muuttuu raja-arvojen alittuessa. Taustalla signaali tunnetaan endplate potentiaali (EPP), koska se on mahdollinen muutos kirjataan moottori endplate. Yleisesti tiedetään, eksitatoristen postsynaptisten potentiaali (EPSP).,
Kuraren estää endplate potentiaalia, koska se on kilpailukykyinen estäjä asetyylikoliini (ACh), lähetin vapautetaan presynaptinen pääte. Kuraren ei estä jännite-riippuvaisten Na+ johtokyky tai jännite-riippuvainen K+ johtokyky, että taustalla lihaksen aktiopotentiaalin. Curare vaikuttaa ärsykkeeseen (EPSP), joka yleensä johtaa lihasten vaikutuspotentiaalin aloittamiseen. Eläin, joka on myrkytetty kuraren tukehtuvat koska prosessi hermolihasliitokseen klo hengitysteiden lihaksia on estetty.,
normaalisti päätepotentiaalin suuruus on melko suuri. Itse asiassa päätepotentiaalin amplitudi on noin 50 mV, mutta kynnyksen saavuttamiseen tarvitaan vain noin 30 mV. Ylimääräistä 20 mV: tä kutsutaan turvallisuustekijäksi. Näin ollen, vaikka endplate potentiaalia oli tullut jonkin verran pienempi (esim., 40 mV amplitudi), koska väsymys, EPP olisi reach-asetuksen kynnys-ja one-to-one suhde välillä aktiopotentiaalin moottorin axon ja aktiopotentiaalin lihas solu olisi säilynyt.
Kuva 4.4
4.,3 Eteneminen EPP –
Mitkä ovat ominaisuuksia EPP ja miten se verrata ominaisuuksia aktiopotentiaalin?
On endplate mahdollinen, koska jännite-riippuvainen muutos Na+ ja K+ permeabilities kuten aktiopotentiaalin?
Onko endplate-potentiaali propagoitu toimintapotentiaalin kaltaisella tavalla?
vasemmalla oleva luku havainnollistaa koetta, jossa tutkitaan endplaattipotentiaalin leviämistä. Lihaskuitu lävistetään toistuvasti elektrodeilla 1 mm välein., (Huomaa, että endplate potentiaali on pieni, koska tämä koe on valmis, kun läsnä on alhainen pitoisuus kuraren joten endplate potentiaalia voidaan tallentaa ilman komplikaatioita, se voi aiheuttaa aktiopotentiaalin.) Endplate-potentiaalia ei levitetä kokonaan tai ei lainkaan. Se kyllä leviää lihaksen mukana, mutta se tekee niin säädyllisesti., Näin ollen leviäminen endplate mahdollinen sen päällä aloittamisesta ja muita sivustoja sekä lihassolujen tapahtuu passiivisesti ja dekrementoidaan, aivan kuten raja-arvojen alittuessa mahdollinen muutos yhdessä osa axon leviää pitkin axon, tai vain koska lämpötilan muutos jossain vaiheessa metallinen tanko leviää pitkin sauva.
4.4 Yhteenveto tapahtumaketju Taustalla EPP
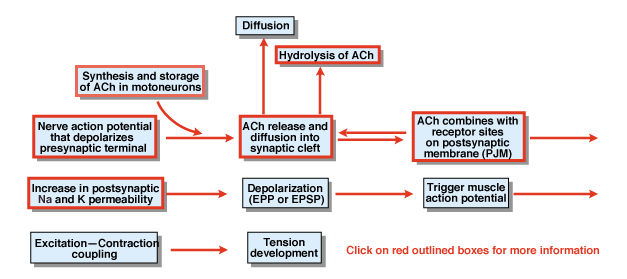
Kuvassa 4.5,
Mitkä ovat muut prosessin vaiheet kemiallisten synaptic siirto? Kuva 4.,5 tarjoaa yleiskuvan. Hermo aktiopotentiaalin, joka on aloitettu solun kehon selkärangan moottori neuroni etenee ulos vatsanpuoleinen juuret ja lopulta tunkeutuu synaptisten terminaalien moottori neuronien. Seurauksena aktiopotentiaalin, kemiallinen lähetin asetyylikoliini (ACh) vapautuu synapsirakoon. ACh diffundoituu synaptisen kleftin poikki ja sitoutuu postsynaptisen tai postjunktionaalisen kalvon erityisiin reseptoreihin. Sitova ACh sen reseptoreihin tuottaa conformational muuttaa kalvo kanava, joka on erityisesti läpäisevä sekä Na+ ja K+., Na+ – ja K+ – läpäisevyyden lisääntymisen seurauksena postsynaptinen kalvo depolarisoituu. Että depolarisaatio on nimeltään endplate mahdollisia tai yleisemmin EPSP. Jos EPSP on riittävän suuri, koska se yleensä on hermo-lihasliitoksessa, se johtaa aloittamisen aktiopotentiaalin lihas solu. Toimintapotentiaali käynnistää herätesupistuskytkimen prosessin ja jännityksen kehittymisen. Endplate-potentiaalin kesto on noin 10 msek.
kaksi tekijää ohjaavat EPSP: n kestoa hermoliitoksessa., Ensin ACh poistetaan diffuusiolla. Toiseksi, aine synapsiraossa, nimeltä asetyylikoliiniesteraasi (AChE), hydrolysoi tai hajoaa ACh. AChE on yksi tehokkaimmista tunnetuista entsyymeistä. Yksi AChE-molekyyli voi hydrolysoida 600 000 Ach-molekyyliä minuutissa.
4.5 Rooli Särky
Kuva 4.6
tärkeä perhe-aineita, joista yksi on neostigmiini, estää toiminnan Särkee. Neostigmiini estää AChE: n toiminnan ja tekee siten endplate-potentiaalista suuremman ja pidemmän keston., Tämä luku kuvaa kahta päätetapahtumaa. Yksi kirjattiin keittosuolaliuoksessa ja curaressa ja toinen sen jälkeen, kun neostigmiini lisättiin liuokseen. (Kuraren on lisätty niin, että ominaisuudet EPP voidaan tutkia ilman, että aiheutuu aktiopotentiaalin lihas solu.) Neostigmiinin käytön jälkeen endplaattipotentiaali on paljon suurempi ja pitempi.
4.6 on Myasthenia Gravis
Myasthenia gravis liittyy vaikea lihasheikkous, koska lasku määrä asetyylikoliinin reseptoreihin lihassolujen., Jos endplate potentiaali on pienempi, endplate potentiaali eivät pääse kynnys. Jos se ei saavuta kynnystä, lihassolussa ei ole toimintapotentiaalia eikä lihasten supistumista, mikä aiheuttaa lihasheikkoutta. Neostigmiinia ja muita AChE: n estäjiä käytetään myasthenia gravis-potilaiden hoitoon. Nämä aineet tekevät asetyylikoliinin määrä, joka vapautuu tehokkaammin saavuttaa loput asetyylikoliinireseptorit.
4.,6b hermomyrkkyä
Vaikka estäjät Särky on tärkeä terapeuttinen arvo, jotkut estäjät ovat olleet, ja ovat yhä käytetään myrkkyjä. Jotkut Särkynestäjät, kuten Somaani ja sariini, muodostavat melko peruuttamattoman Särkylohkon. Tämä lohko johtaa äärimmäistä ACh synapsiraossa. Näin myrkytetyt yksilöt kuolevat kohtauksiin ja lihasten spastisuuteen, mukaan lukien hengityslihakset.
Kuva 4.7.
4.,7 Iontoforeesi ACh
Iontoforeesi on mielenkiintoinen tekniikka, jota voidaan edelleen testata hypoteesia, että ACh on välittäjäaine, aine hermo-lihasliitoksessa. Jos ACh-on-lähetin, joka on julkaissut tämän synapse, voisi ennustaa, että sen pitäisi olla mahdollista korvata keinotekoinen sovellus lähettimen normaalin vapauta lähettimen. Koska ACh on positiivisesti varautunut molekyyli, se voidaan pakottaa ulos mikroelektrodi simuloida vapauttaa ACh alkaen presynaptinen pääte.
Kuva 4.,8
Todellakin, minuutti määriä ACh voidaan soveltaa muun muassa lihasliitos. Kuva 4.8 verrataan lisäksi EPP tuotettu stimulaation moottori axon ja vastaus ulos paiskautuminen ACh. Mahdollinen muutos näyttää lähes identtiseltä ACh: n normaalin vapautumisen tuottaman endplate-potentiaalin kanssa. Tämä kokeilu tarjoaa kokeellista tukea käsitteelle, että ACh on luonnollinen lähetin tässä synapse.,
vastaus poisto ACh on joitakin muita mielenkiintoisia ominaisuuksia, jotka kaikki ovat sopusoinnussa kolinergisen luonne synapse klo luuston lihasliitos. Neostigmiini tekee ACh: n iontoforeesille vasteen pidemmäksi ja suuremmaksi. Curare vähentää vastetta, koska se kilpailee ACh: n normaalin sitoutumisen kanssa. Jos ACh työnnetään ulos lihassolujen, mitään ei tapahdu koska reseptorit asetyylikoliinin eivät sisällä; he ovat ulkopuolella lihassolujen., Soveltaminen asetyylikoliinin alueiden lihasten pois end-levy tuottaa mitään vastausta, koska reseptoreihin ACh ovat keskittyneet tällä synaptic alueella.
testaa ymmärrystä niin pitkälle, pohtia, miten agentti, kuten TTX vaikuttaisi sukupolven sekä EPP ja vaste lihaksen kuitua iontophoretic sovellus ACh? TTX: llä ei ole vaikutusta ACh: n vasteeseen, mutta se estää EPP: n. Syy vastaus ACh ei muutu, on selvää, mutta monet odottavat, että jos on mitään vaikutusta tässä ei pitäisi olla mitään vaikutusta EPP joko., Tetrodotoksiini ei vaikuta sitova asetyylikoliini reseptoreihin, ja näin ollen ei vaikuta vastaus suora soveltaminen ACh. Tetrodotoksiini vaikuttaa kuitenkin vaikutuspotentiaalin syntymiseen moottoriaksonissa. Jos aktiopotentiaalia ei voida saada aikaan moottoriaksonissa, se ei voi aiheuttaa lähettimen vapautumista. Näin tetrodotoksiini poistaisi EPP: n kokonaan. Lohko ei olisi, koska lohko ACh-reseptoreihin, vaan pikemminkin estää joitakin vaihe ennen vapauttamista lähettimen.
4.,8 Ionic Mekanismeja EPP –
Bernard Katz ja hänen kollegansa olivat edelläkävijöitä tutkitaan mekanismeja synaptic siirto hermo-lihasliitoksessa. He ehdottivat, että ACh: n avaama kanava olisi yhtä läpäisevä kuin sekä Na+ että K+. Koska se oli yhtä läpäisevä Na+ ja K+, Katz ehdotti, että seurauksena avaaminen näitä kanavia, kalvon potentiaalia olisi siirtyä kohti 0 mV. (Alfan arvo GHK-yhtälössä on yhtä suuri, joka yhtälöön korvattuna tuottaa noin 0 mV: n potentiaalin.,)
Kuva 4.9
kokeilu kuvassa vasemmalla testit käsite. Lihassolujen on tunkeutunut tallennus elektrodi sekä toinen elektrodi, joka voidaan liittää sopiva lähde potentiaalia, jotta voidaan keinotekoisesti muuttaa solukalvon potentiaalia. Normaalisti kalvopotentiaali on taas noin -80 mV, pieni määrä curarea lisätään niin, että EPP on pieni. Katz huomasi näissä kokeissa, että EPP: n koko muuttui dramaattisesti riippuen lihassolun potentiaalista., Jos kalvopotentiaali siirretään 0 mV: hen, potentiaalista muutosta ei kirjata lainkaan. Jos kalvopotentiaalia tehdään +30 mV, EPP käännetään ylösalaisin. Niinpä kolme erilaista ärsykettä tuottavat endplate-potentiaaleja, jotka ovat hyvin erilaisia toisistaan.
puute vastaus, kun potentiaali on 0 mV on erityisen informatiivinen. Mieti, miksi mahdollista muutosta ei kirjata. Oletettavasti lähetin vapautuu ja sitoutuu reseptoreihin., Yksinkertainen selitys mahdollisen muutoksen puutteelle on se, että mahdollisuudet, joihin ACh-kanavien avaaminen pyrkii, on jo saavutettu. Jos kalvopotentiaali saadaan positiivisemmaksi kuin 0 mV, EPP käännetään ylösalaisin. Olipa potentiaali mikä tahansa, läpäisevyyden muutos pyrkii siirtämään kalvopotentiaalia kohti 0 mV! Jos lepopotentiaali on enemmän negatiivinen kuin 0 mV, on ylöspäin suuntautuva taipuma. Jos se on positiivisempi, edessä on alaspäin suuntautuva taipuma. Jos se on jo 0 mV, ei ole taipuma.
Kuva 4.,10
Tämä potentiaali on myös nimeltään kääntyminen mahdollinen, koska se on potentiaali, jolla merkki synaptinen potentiaali kääntää. Kokeilu osoittaa, että seurauksena ACh sitovia reseptoreita, erityisiä kanavia tullut yhtä läpäisevä Na+ ja K+. Tämä läpäisevyysmuutos pyrkii siirtämään kalvopotentiaalia sieltä, missä se aluksi on, kohti uutta 0 mV: n potentiaalia.
Miksi normaali endplate mahdollisten koskaan todella saavuttaa 0 mV?, Yksi syy on, että sekvenssi läpäisevyys muuttuu, että taustalla aktiopotentiaalin ”suolla ulos” muutokset valmistettu EPP. EPP ei kuitenkaan yltäisi 0 mV: hen, vaikka toimintapotentiaalia ei käynnistettäisikään. Tämä johtuu siitä, että ACh-kanavat ovat vain pieni osa lihasten kuiduissa olevien kanavien kokonaismäärästä. Myös K + – kanavat, jotka tuottavat lihassolun lepopotentiaaleineen, ovat läsnä. Heidän tehtävänään on yrittää pitää solu lepopotentiaalissa.,
kanava avata ACh on jäsenenä yleisen luokan kanavia kutsutaan ligandi-aidatulla kanavia tai ionotropic reseptoreihin. Kuten kuvassa 4.10 esitetään, lähettimen sidontapaikka on osa itse kanavaa. Seurauksena lähetin sitova reseptorin (yleensä kaksi molekyyliä ovat tarpeen), on conformational muuttaa proteiinin jolloin huokosten alueella avata ja ionit virtaavat alas heidän sähkökemiallinen kaltevuudet. Kanavan tarkemmat tiedot esitetään luvussa 11.,
Testaa tietosi
- Kysymys 1
- A
- B
- C
- D
On endplate potentiaalia lihasten solu voisi periaatteessa olla valmistettu vähentynyt läpäisevyys mikä seuraavista ioneja(s)? (Olettaa, että siellä on rajallinen alkuperäinen läpäisevyys kunkin ioneja alla ja että fysiologinen pitoisuus kaltevuudet ovat läsnä.):
A. Na+
B. Na+ ja Ca2+
C Ca2+
D., K+
On endplate potentiaalia lihasten solu voisi periaatteessa olla valmistettu vähentynyt läpäisevyys mikä seuraavista ioneja(s)? (Olettaa, että siellä on rajallinen alkuperäinen läpäisevyys kunkin ioneja alla ja että fysiologinen pitoisuus kaltevuudet ovat läsnä.):
A. Na+ tämä vastaus on virheellinen.
end-levy potentiaalia on depolarisaatio, joka on yleensä valmistettu samanaikainen kasvu läpäisevyys natrium-ja kalium-ioneja., Jos natriumin läpäisevyys vähenisi selektiivisesti, tällainen väheneminen ei johtaisi depolarisaatioon. Pikemminkin se johtaisi itse asiassa hyperpolarisaatioon, koska alfa GHK-yhtälössä vähenisi. Suhde läpäisevyys natrium-ja kalium olisi suosinut kohti kaliumin läpäisevyyttä, liikkuvat kalvon potentiaalia kohti kalium tasapainon potentiaalia ja tuottaa hyperpolarization.
B. Na+ ja Ca2+
C Ca2+
D., K+
On endplate potentiaalia lihasten solu voisi periaatteessa olla valmistettu vähentynyt läpäisevyys mikä seuraavista ioneja(s)? (Olettaa, että siellä on rajallinen alkuperäinen läpäisevyys kunkin ioneja alla ja että fysiologinen pitoisuus kaltevuudet ovat läsnä.):
A. Na+
B. Na+ – ja Ca2+ – Tämä vastaus on VÄÄRÄ.
Jos läpäisevyys natrium-ja kalsium-tableteista pieneni, niiden seuraukset olisivat samanlaisia, että valinta A. Vähentämällä natriumin läpäisevyys yksin olisi yleensä hyperpolarize solun., Vastaavasti kalsiumin läpäisevyyden vähentäminen saattaa myös hyperpolarisoida solun. Kalsiumin tasapainon potentiaali on hyvin positiivinen arvo, ja jos siellä oli joitakin tonic lepää läpäisevyys kalsiumia, joka läpäisevyys edistää tonic depolarisaatio kalvon potentiaalia. Näin ollen kalsiumin läpäisevyyden väheneminen poistaisi tuon toonisen depolarisoivan vaikutuksen ja johtaisi hyperpolarisaatioon.
C. Ca2 +
D., K+
On endplate potentiaalia lihasten solu voisi periaatteessa olla valmistettu vähentynyt läpäisevyys mikä seuraavista ioneja(s)? (Olettaa, että siellä on rajallinen alkuperäinen läpäisevyys kunkin ioneja alla ja että fysiologinen pitoisuus kaltevuudet ovat läsnä.):
A. Na+
B. Na+ ja Ca2+
C Ca2+ Tämä vastaus on VÄÄRÄ.
pelkän kalsiumin läpäisevyyden väheneminen aiheuttaisi, jos mitään, hyperpolarisaation. KS. logic of response to choice B.
D., K+
On endplate potentiaalia lihasten solu voisi periaatteessa olla valmistettu vähentynyt läpäisevyys mikä seuraavista ioneja(s)? (Olettaa, että siellä on rajallinen alkuperäinen läpäisevyys kunkin ioneja alla ja että fysiologinen pitoisuus kaltevuudet ovat läsnä.):
A. Na+
B. Na+ ja Ca2+
C Ca2+
D. K+ Tämä vastaus on OIKEA!
lasku kaliumin läpäisevyyttä johtaisi depolarisaatio samanlainen loppu-levy potentiaalia., Tämä johtuu siitä, että on levossa tonic läpäisevyys kalium ja natrium. Suuri kaliumin läpäisevyys pyrkii pitämään kalvopotentiaalin lähellä kaliumin tasapainopotentiaalia. Jos se lepää läpäisevyys on vähentynyt, alfa Goldman yhtälö olisi tullut suurempi arvo, liikkuvat kalvon potentiaalia hieman lähempänä natrium-tasapainon mahdollinen (eli depolarisaatio).