Figur 4.1 (se forstørret visning)
Derfor, at en forståelse af denne synapse fører til en forståelse af de andre. Derfor vil vi først diskutere processen med synaptisk transmission ved skeletets neuromuskulære kryds.,
funktionerne i det synaptiske kryds ved det neuromuskulære kryds er vist i figuren til venstre. Skeletmuskelfibre er inderveret af motorneuroner, hvis cellelegemer er placeret i rygmarvets ventrale horn. Den terminale region af aononen giver anledning til meget fine processer, der løber langs skeletmuskelceller. Langs disse processer er specialiserede strukturer kendt som synapser. Den særlige synapse, der er lavet mellem en spinalmotorisk neuron og skeletmuskelcelle, kaldes den motoriske endeplade på grund af dens specifikke struktur.,
synapsen ved det neuromuskulære kryds har tre karakteristiske træk ved kemiske synapser i nervesystemet. For det første er der en tydelig adskillelse mellem den presynaptiske og den postsynaptiske membran. Mellemrummet mellem de to er kendt som den synaptiske kløft. Rummet fortæller os, at der skal være en vis mellemliggende signalmekanisme mellem den presynaptiske neuron og den postsynaptiske neuron for at få informationsstrøm over den synaptiske kløft. For det andet er der en karakteristisk høj densitet af små sfæriske vesikler., Disse synaptiske vesikler indeholder neurotransmitterstoffer. Synapser er også forbundet med en høj densitet af mitokondrier. For det tredje, i de fleste tilfælde, at der er karakteristisk fortykkelse af den postsynaptiske membran, som skyldes ikke mindst det faktum, at den postsynaptiske membran har en høj tæthed af specialiserede receptorer, der binder kemisk transmitter stoffer, der frigives fra den presynaptic neuron. Yderligere detaljer om de morfologiske træk ved synaptiske kryds findes i kapitel 8 og Kapitel 10.
4.,2 Fysiologi af Synaptisk Transmission i den Neuromuskulære Junction
Figur 4.2
figuren til højre illustrerer på en meget skematisk måde, hvordan det er muligt at studere fysiologi af synaptisk transmission på skelet neuromuskulære junction i stor detalje. Et stykke muskel og dets vedhæftede nerve placeres i et lille eksperimentelt kammer fyldt med en passende Ringeopløsning. Muskelcellens hvilepotentiale registreres med en mikroelektrode. Elektroder placeres også på overfladen af nerveaksonet., Korte elektriske stød forårsager handlingspotentialer, der skal initieres, som forplantes til den synaptiske terminal.
figuren nedenfor illustrerer to typer potentielle ændringer, der blev registreret i et sådant isoleret nerve-muskelpræparat. Eksperimentet illustrerer også egenskaberne af et stærkt lægemiddel, curare, som har vist sig at være meget nyttigt til at studere processen med synaptisk transmission ved skeletets neuromuskulære kryds. Del A illustrerer sekvensen af potentielle ændringer registreret i muskelcellen som et resultat af stimulering af motoraksonet., Pilen angiver det tidspunkt, hvor stød leveres til motoraksonet. Bemærk, at der er en hvilende periode efter chokket. Forsinkelsen skyldes den tid det tager for handlingspotentialet i motoraksonet at formere sig fra dets initieringssted. Efter forsinkelsen er der to typer potentialer registreret i muskelcellen. For det første er der et relativt langsomt skiftende potentiale, der vil være i fokus i den følgende diskussion., Hvis det langsomme startpotentiale er tilstrækkeligt stort, som det normalt er i skeletmuskelceller, fremkaldes et andet potentiale, et handlingspotentiale, i muskelcellen.
figur 4.3
handlingspotentialer i skeletmuskelceller skyldes Ioniske mekanismer, der ligner dem, der er diskuteret tidligere. Specifikt er der en spændingsafhængig ændring i Na+ permeabilitet efterfulgt af en forsinket stigning i K+ permeabilitet. (For glatte muskelceller og hjertemuskelceller er de ioniske mekanismer imidlertid forskellige.,)
den underliggende begivenhed, der udløser handlingspotentialet, kan afsløres ved at drage fordel af curare, en pilegift, der bruges af nogle sydamerikanske indianere. En lav dosis curare (del B) reducerer den underliggende begivenhed, men den er stadig ikke tilstrækkeligt reduceret til at falde under tærsklen. Hvis en noget højere dosis curare leveres (del C), bliver den langsomme underliggende begivenhed subthreshold. Det underliggende signal er kendt som endepladepotentialet (EPP), fordi det er en potentiel ændring registreret ved motorens endeplade. Generelt er det kendt som et e .citatorisk postsynaptisk potentiale (EPSP).,
Curare blokerer endepladepotentialet, fordi det er en konkurrencedygtig hæmmer af acetylcholin (ACh), transmitteren frigivet ved den presynaptiske terminal. Curare blokerer ikke den spændingsafhængige Na+ konduktans eller den spændingsafhængige K+ konduktans, der ligger til grund for muskelvirkningspotentialet. Curare påvirker stimulus (EPSP), som normalt fører til initiering af muskelvirkningspotentialet. Et dyr, der er forgiftet med curare, vil kvæles, fordi processen med neuromuskulær transmission ved respiratoriske muskler er blokeret.,
normalt er størrelsen af endepladepotentialet ret stort. Faktisk er amplituden af endepladepotentialet omkring 50 mV, men kun omkring 30 mV er nødvendig for at nå tærskelværdien. Den ekstra 20 mV kaldes sikkerhedsfaktoren. 40 mV i amplitude) på grund af træthed, ville EPP nå tærsklen, og en-til-en-forholdet mellem et handlingspotentiale i motoraksonet og et handlingspotentiale i muskelcellen ville blive bevaret.
Figur 4.4
4.,3 udbredelse af EPP
Hvad er EPP ‘ s egenskaber, og hvordan sammenlignes det med handlingspotentialets egenskaber?
er endepladepotentialet på grund af en spændingsafhængig ændring i na+ og K+ permeabilities som handlingspotentialet?
er endepladepotentialet forplantet på en alt-eller-intet måde som handlingspotentialet?
figuren til venstre illustrerer et eksperiment, der undersøger udbredelsen af endepladepotentialet. Muskelfiberen impaleres gentagne gange med elektroder med 1 mm intervaller., (Bemærk, at endepladepotentialet er lille, fordi dette eksperiment udføres i nærvær af en lav koncentration af curare, så endepladepotentialet kan registreres uden komplikationer ved at udløse et handlingspotentiale.) Endepladen potentiale ikke formeres i en alt-eller-intet måde. Det spredes langs musklen, men det gør det med reduktion., Således forekommer spredningen af endepladepotentialet fra dets initieringssted til andre steder langs muskelcellen passivt og med reduktion, ligesom en subthresholdspotentiale ændring i en del af A .onen spreder sig langs a .onen, eller ligesom en temperaturændring på et tidspunkt på en metalstang spreder sig langs stangen.
4.4 Oversigt over den Sekvens af Begivenheder, der ligger bag EPP
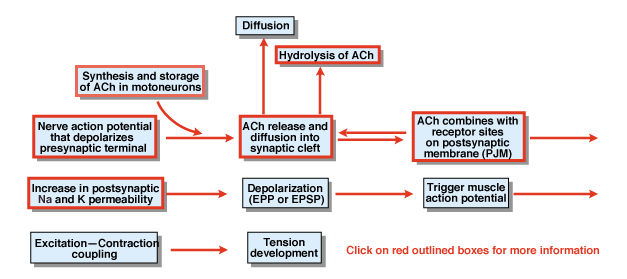
Figur 4.5
Hvad er det andet skridt i processen af kemisk synaptisk transmission? Figur 4.,5 giver et overblik. Et nerveaktionspotentiale, der initieres i cellelegemet i en spinalmotorneuron, udbreder de ventrale rødder og invaderer til sidst de synaptiske terminaler i de motoriske neuroner. Som et resultat af handlingspotentialet frigives den kemiske sender acetylcholin (ACh) i det synaptiske spalte. ACh diffunderer over det synaptiske spalte og binder til specielle receptorer på den postsynaptiske eller den postjunktive membran. Bindingen af ACh til dets receptorer frembringer en konformationsændring i en membrankanal, der specifikt er permeabel for både Na+ og K+., Som følge af en stigning i Na + og K+ permeabilitet er der en depolarisering af den postsynaptiske membran. At depolarisering kaldes endepladen potentiale eller mere generelt EPSP. Hvis EPSP er tilstrækkelig stor, som den normalt er ved det neuromuskulære kryds, fører det til initiering af et handlingspotentiale i muskelcellen. Handlingspotentialet initierer processen med e .citationskontraktionskobling og udvikling af spænding. Varigheden af endepladen potentiale er omkring 10 MSEK.
to faktorer styrer varigheden af EPSP ved det neuromuskulære kryds., For det første fjernes ACh ved diffusion. For det andet hydrolyserer eller nedbryder et stof i den synaptiske kløft, kaldet acetylcholinesterase (AChE). AChE er et af de mest effektive en .ymer, der er kendt. Et enkelt Smertemolekyle kan hydrolysere 600.000 ACH-molekyler pr.
4.5 Rolle for Smerte
Figur 4.6
En vigtig gruppe af stoffer, hvoraf den ene er neostigmin, hæmmer virkningen af Smerte. Neostigmin blokerer virkningen af AChE, og derved gør endepladen potentiale større og længere varighed., Dette tal illustrerer to endeplade potentialer. Den ene blev optaget i saltvand og curare og en anden registreret efter neostigmin blev tilsat til opløsningen. (Curare tilføjes, så EPP ‘ s egenskaber kan studeres uden at udløse et handlingspotentiale i muskelcellen.) Efter påføring af neostigmin er endepladepotentialet meget større og længere. 4.6 a Myasthenia Gravis
Myasthenia gravis er forbundet med svær muskelsvaghed på grund af et fald i antallet af acetylcholinreceptorer i muskelcellen., Hvis endepladepotentialet er mindre, vil endepladepotentialet ikke nå tærskelværdien. Hvis det ikke når tærsklen, vil der ikke være noget handlingspotentiale i muskelcellen og ingen sammentrækning af muskelen, hvilket forårsager muskelsvaghed. Neostigmin og andre hæmmere af AChE bruges til behandling af patienter med myasthenia gravis. Disse midler gør mængden af acetylcholin, der frigives mere effektivt, til de resterende acetylcholinreceptorer.
4.,6b nervemidler
selvom hæmmere af AChE har vigtig terapeutisk værdi, har nogle hæmmere været og bruges stadig som giftstoffer. Nogle AChE hæmmere såsom Soman og Sarin danner en temmelig irreversibel blok af smerte. Denne blok fører til ekstreme niveauer af ACh i den synaptiske kløft. Personer, der er så forgiftet, dør af anfald og muskelspasticitet, herunder åndedrætsmuskler.
Figur 4.7
4.,7 iontoforese af ACH
iontoforese er en interessant teknik, der kan bruges til yderligere at teste hypotesen om, at ACH er neurotransmitterstoffet ved det neuromuskulære kryds. Hvis ACh er senderen, der frigives af denne synapse, ville man forudsige, at det skulle være muligt at erstatte kunstig anvendelse af senderen til normal frigivelse af senderen. Da ACh er et positivt ladet molekyle, kan det tvinges ud af en mikroelektrode for at simulere frigivelsen af ACH fra en presynaptisk terminal.
Figur 4.,8
faktisk kan små mængder ACh påføres i nærheden af det neuromuskulære kryds. Figur 4.8 sammenligner en EPP produceret ved stimulering af motorakson og respons på udstødninger af ACh. Den potentielle ændring ser næsten identisk med endepladepotentialet produceret ved den normale frigivelse af ACH. Dette eksperiment giver eksperimentel støtte til konceptet om, at ACH er den naturlige sender ved denne synapse.,
svaret på udstødningen af ACh har nogle andre interessante egenskaber, som alle er i overensstemmelse med den kolinergiske natur af synapsen ved skeletets neuromuskulære kryds. Neostigmin gør responsen på iontoforese af ACh længere og større. Curare reducerer svaret, fordi det konkurrerer med den normale binding af ACH. Hvis ACh skubbes ud i muskelcellen, sker der intet, fordi receptorerne for acetylcholin ikke er inde; de er på ydersiden af muskelcellen., Anvendelse af acetylcholin til områder af musklen væk fra endepladen producerer intet Svar, fordi receptorerne for ACh er koncentreret i det synaptiske område.
for at teste din forståelse hidtil skal du overveje, hvordan en agent som TT?ville påvirke genereringen af både en EPP og responsen af en muskelfiber til den iontoforetiske anvendelse af ACH? TT.har ingen effekt på svaret på ACh, men det blokerer EPP. Årsagen til, at reaktionen på ACh er upåvirket, er klar, men mange forventer, at hvis der ikke er nogen effekt her, bør der heller ikke være nogen effekt på PPE., Tetrodoto .in påvirker ikke bindingen af acetylcholin til receptorerne og påvirker derfor ikke responsen på direkte anvendelse af ACH. Tetrodoto .in vil imidlertid påvirke evnen af et handlingspotentiale, der skal fremkaldes i motoraksonet. Hvis et handlingspotentiale ikke kan fremkaldes i motoraksonet, kan det ikke forårsage frigivelse af senderen. Tetrodoto .in ville således helt afskaffe EPP. Blokken skyldes ikke en blok af ACh-receptorer, men snarere en blok af et trin før frigivelsen af senderen.
4.,8 Ioniske mekanismer i EPP
Bernard kat.og hans kolleger var pionerer i at undersøge mekanismer for synaptisk transmission ved det neuromuskulære kryds. De foreslog, at kanalen åbnet af ACh var en, der havde lige permeabilitet til både Na + og K+. Fordi det var lige så gennemtrængeligt for Na+ og K+, foreslog kat., at membranpotentialet som følge af åbningen af disse kanaler ville bevæge sig mod 0 mV. (En værdi af alfa i GHK ligning lig med en, som når substitueret i ligningen, giver et potentiale på ca. 0 mV.,)
figur 4.9
eksperimentet vist i figuren til venstre tester dette koncept. Muskelcellen er blevet trængt ind med en optageelektrode såvel som en anden elektrode, der kan forbindes til en passende potentialekilde for kunstigt at ændre membranpotentialet. Normalt er membranpotentialet omkring -80 mV igen, en lille mængde curare tilsættes, så EPP er lille. Kat.bemærkede i disse eksperimenter, at EPP ‘ s størrelse ændrede sig dramatisk afhængigt af muskelcellens potentiale., Hvis membranpotentialet flyttes til 0 mV, registreres ingen potentiel ændring overhovedet. Hvis membranpotentialet er lavet + 30 mV, er EPP omvendt. Så tre forskellige stimuli producerer endepladepotentialer, der er meget forskellige fra hinanden.
manglen på et svar, når potentialet er ved 0 mV, er særlig informativ. Overvej hvorfor der ikke registreres nogen potentiel ændring. Formentlig bliver transmitteren frigivet og bundet til receptorerne., Den enkle forklaring på manglende potentiel ændring er, at det potentiale, som åbningen af ACh-kanaler forsøger at nå, allerede er opnået. Hvis membranpotentialet gøres mere positivt end 0 mV, er EPP omvendt. Uanset potentialet har ændringen i permeabilitet tendens til at bevæge membranpotentialet mod 0 mV! Hvis hvilepotentialet er mere negativt end 0 mV, er der en opadgående afbøjning. Hvis det er mere positivt, er der en nedadgående afbøjning. Hvis det allerede er på 0 mV, er der ingen afbøjning.
Figur 4.,10
dette potentiale kaldes også reverseringspotentialet, fordi det er det potentiale, hvor tegnet på det synaptiske potentiale vender tilbage. Eksperimentet indikerer, at specifikke kanaler som følge af ACH-binding til receptorer bliver lige gennemtrængelige for Na+ og K+. Denne permeabilitetsændring har en tendens til at bevæge membranpotentialet, uanset hvor det oprindeligt er mod et nyt potentiale på 0 mV.
hvorfor når det normale endepladepotentiale aldrig 0 mV?, En af grundene er, at sekvensen af permeabilitetsændringer, der ligger til grund for handlingspotentialet “sump ud” de ændringer, der produceres af EPP. Men selv hvis et handlingspotentiale ikke blev udløst, ville EPP stadig ikke nå 0 mV. Dette skyldes, at ACH-kanalerne kun er en lille brøkdel af det samlede antal kanaler i muskelfibre. K + – kanalerne, der giver en muskelcelle med sit hvilepotentiale, er også til stede. Deres job er at forsøge at opretholde cellen ved hvilepotentialet., den kanal, der åbnes af ACh, er medlem af en generel klasse af kanaler kaldet ligand-gatede kanaler eller ionotrope receptorer. Som illustreret i figur 4.10 er senderens bindingssted en del af selve kanalen. Som et resultat af transmitterbinding til receptoren (generelt er to molekyler nødvendige), der er en konformationsændring i proteinet, der tillader et poreområde at åbne og ioner at strømme ned ad deres elektrokemiske gradienter. Yderligere oplysninger om kanalen er præsenteret i kapitel 11.,
Test Din Viden
- punkt 1
- En
- B
- C
- D
En endplate potentiale i en skeletal muscle cell kunne i princippet være produceret af en nedsat permeabilitet, hvilke af de følgende ioner(s)? (Antag, at der er en endelig indledende permeabilitet for hver af de ioner, der er anført nedenfor, og at fysiologiske koncentrationsgradienter er til stede.):
A. Na+
B. Na+og Ca2 +
C. Ca2 +
D., K +
et endepladepotentiale i en skeletmuskelcelle kunne i princippet produceres ved en nedsat permeabilitet til hvilken af følgende ioner? (Antag, at der er en endelig indledende permeabilitet for hver af de ioner, der er anført nedenfor, og at fysiologiske koncentrationsgradienter er til stede.):
A. Na+ dette svar er forkert.
et endepladepotentiale er en depolarisering, der normalt produceres ved den samtidige stigning i permeabiliteten for natrium-og kaliumioner., Hvis der var et selektivt fald i natriumpermeabilitet, ville et sådant fald ikke føre til en depolarisering. Snarere ville det faktisk føre til en hyperpolarisering, fordi alfa i GHK-ligningen ville blive reduceret. Forholdet mellem permeabilitet til natrium og kalium ville være begunstiget i retning af kaliumpermeabilitet, bevæger membranen potentiale i retning af kalium ligevægt potentiale og producerer en hyperpolarisering.
B. Na+ og Ca2+
C. Ca2 +
D., K +
et endepladepotentiale i en skeletmuskelcelle kunne i princippet produceres ved en nedsat permeabilitet til hvilken af følgende ioner? (Antag, at der er en endelig indledende permeabilitet for hver af de ioner, der er anført nedenfor, og at fysiologiske koncentrationsgradienter er til stede.):
A. Na+
B. Na+ og Ca2+ dette svar er forkert.
Hvis permeabiliteten for natrium og calcium blev nedsat, ville deres konsekvenser svare til den i valg A. nedsættelse af natriumpermeabiliteten alene ville have tendens til at hyperpolarisere cellen., Tilsvarende kan nedsættelse af permeabiliteten for calcium også hyperpolarisere cellen. Calcium ligevægtspotentialet er en meget positiv værdi, og hvis der var noget tonisk hvilepermeabilitet for calcium, ville denne permeabilitet bidrage til en tonisk depolarisering af membranpotentialet. Derfor ville et fald i calciumpermeabiliteten fjerne den toniske depolariserende virkning og resultere i en hyperpolarisering.
C. Ca2 +
D., K +
et endepladepotentiale i en skeletmuskelcelle kunne i princippet produceres ved en nedsat permeabilitet til hvilken af følgende ioner? (Antag, at der er en endelig indledende permeabilitet for hver af de ioner, der er anført nedenfor, og at fysiologiske koncentrationsgradienter er til stede.):
A. Na+
B. Na+ og Ca2+
C. Ca2+ dette svar er forkert.
et fald i calciumpermeabiliteten alene ville, hvis noget, producere en hyperpolarisering. Se logik for respons på valg B.
D., K +
et endepladepotentiale i en skeletmuskelcelle kunne i princippet produceres ved en nedsat permeabilitet til hvilken af følgende ioner? (Antag, at der er en endelig indledende permeabilitet for hver af de ioner, der er anført nedenfor, og at fysiologiske koncentrationsgradienter er til stede.):
A. Na+
B. Na+ og Ca2+
C. Ca2+
D. K+ dette svar er korrekt!
et fald i kaliumpermeabiliteten ville føre til en depolarisering svarende til et endepladepotentiale., Dette skyldes, at der i ro er en tonisk permeabilitet for kalium og natrium. Den høje permeabilitet for kalium har en tendens til at holde membranpotentialet nær kaliumbalancepotentialet. Hvis denne hvilepermeabilitet reduceres, vil alfa i Goldman-ligningen blive en større værdi, der bevæger membranpotentialet lidt tættere på natriumbalancepotentialet (dvs.en depolarisering).