Indledning
Darwins stor indsigt, og det samlende princip for biologi i dag, er, at alle arter er relateret til hinanden som søstre, fætre og kusiner, og fjerne slægtninge i et stort family tree of life. Implikationerne er betagende; hvis vi kunne rejse langt nok tilbage i tiden, ville vi finde fælles forfædre mellem os selv og enhver anden levende organisme, fra pindsvin til flamingoer til kaktus., Vores umiddelbare evolutionære familie består af hominoiderne, gruppen af primater, der inkluderer de ” mindre aber “(siamangs og gibbons) såvel som de “store aber” (chimpanser, bonoboer, gorillaer og orangutanger). Blandt de store aber er vores nærmeste pårørende chimpanser og bonoboer (Figur 1). Fossilrekorden sammen med undersøgelser af menneske-og ape-DNA indikerer, at mennesker delte en fælles stamfar med chimpanser og bonoboer engang for omkring 6 millioner år siden (mya)., Vi begynder denne diskussion af vores Arts udvikling i Afrika, nær slutningen af den geologiske tidsperiode kendt som Miocene, lige før vores afstamning afveg fra chimpanser og bonobos.
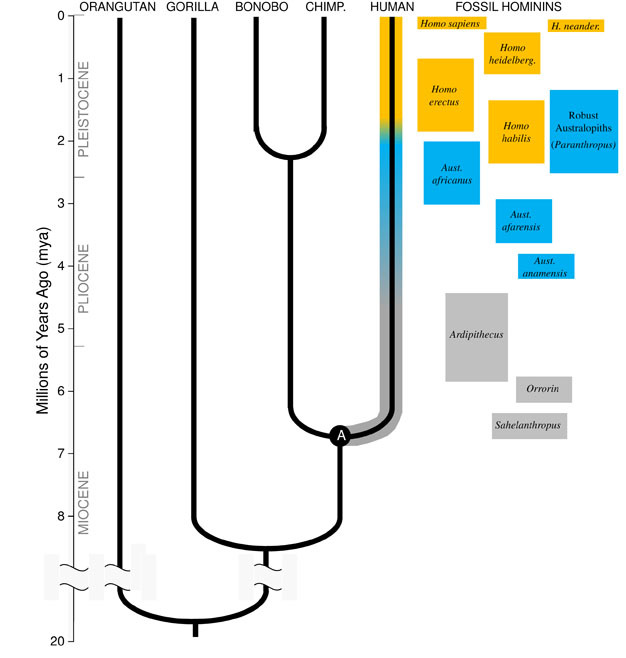
Miocæn Oprindelsen af Hominin Slægt
for at forstå udviklingen af enhver art, må vi først fastlægge dens forfædres tilstand: hvad slags dyr, udviklede det sig fra? For vores afstamning kræver dette, at vi prøver at rekonstruere den sidste fælles stamfar til mennesker og sjimpanser (markeret “a” i Figur 1). Den Human-chimpanse sidste fælles forfader (HC-LCA) er den art, hvorfra hominin-afstamningen og chimpansen & bonobo-afstamning divergerede., Hominins er arter på vores gren af hominoid træet efter opdelingen med chimpanse & bonobo-line, herunder alle de uddøde arter og evolutionære side-grene (Figur 1).
Der var en stor mangfoldighed af abearter i Miocæn, med snesevis af arter kendt fra fossilrekorden i hele Afrika, Europa og Asien. Disse arter varierede i deres anatomi og økologi, og det er ikke klart, hvilken, hvis nogen, af de fossile arter, der hidtil er opdaget, repræsenterer HC-LCA (Kunimatsu et al. 2007; Young and MacLatchy, 2004)., Ikke desto mindre ved vi fra fossile og sammenlignende beviser, at det lignede meget mere levende aber end levende mennesker. HC-LCA ville have haft en abe-størrelse hjerne og krop, med relativt lange arme og fingre og en gribende fod, der gjorde det muligt for den at foder i træerne. Hundetænderne var sandsynligvis store og skarpe, som det ses i flere Miocæn hominoider. Fremdeles vare hjørnetænderne rimeligvis kønsdimorfe, idet hannerne havde meget større hjørnetænder end hunnerne, som det ses blandt de levende store aber og Miocæn-fossiler., Ligesom levende aber ville den have vandret firdoblet (på alle fire), når den var på jorden, og dens kost ville næsten udelukkende have bestået af plantefødevarer, primært frugt og blade.
tidlige homininer
ændringer fra en abelignende anatomi kan ses i hominoidfossiler fra det sene miocen i Afrika. Nogle hominoidarter fra denne periode udviser træk, der er typiske for mennesker, men som ikke ses i de andre levende aber, der fører paleoanthropologer til at udlede, at disse fossiler repræsenterer tidlige medlemmer af hominin-afstamningen., De første menneskelignende træk, der vises i hominin fossil record, er bipedal gang og mindre, stumpe hjørnetænder.
de ældste homininer, der i øjeblikket er kendt, er Sahelanthropus tchadensis fra Tchad (Brunet et al. 2005) og Orrorin tugenensis fra Kenya (Senut et al. 2001). Sahelanthropus, dateret til mellem 6 og 7 mya, er kendt fra en stort set komplet kranium og nogle andre fragmentariske rester. Dens hjernestørrelse, 360cc, ligger inden for det område, der ses i chimpanser, og kraniet har en massiv panderyg, der ligner tykkelse som mandlige gorillaer (Brunet et al. 2005)., Men den position og orientering af foramen magnum, hullet i bunden af kraniet, hvorigennem rygmarven passerer, tyder på, at Sahelanthropus stod og gik bipedally, med sin rygsøjlen holdes i lodret position som moderne mennesker i stedet for vandret som i aber, og andre firbenede (Zollikofer et al. 2005). Orrorin er primært kendt fra postkraniale fossiler, inklusive en delvis lårben. Den proksimale del af lårbenet viser ligheder med moderne menneskers, hvilket antyder, at arten var bipedal (Pickford et al. 2002)., Ingen kranier af Orrorin er blevet genvundet, og derfor er dens kraniale morfologi og hjernestørrelse usikker. I både Orrorin og Sahelanthropus er hundetænderne hos mænd større og mere spidse end hos moderne mennesker, men er små og stumpe sammenlignet med hjørnetænderne hos mandlige aber. Dette antyder, at seksuel dimorfisme hos hunde — og i forlængelse heraf, konkurrence blandt mænd om parring af adgang til kvinder — blev formindsket i disse tidlige homininer sammenlignet med de store aber.
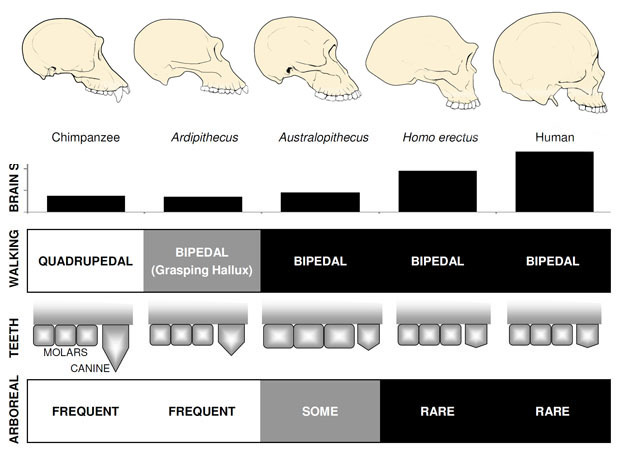
langt den mest kendte tidlige hominin er Ardipithecus ramidus, en 4.,4 millioner år gamle arter fra Etiopien, som er kendt fra et næsten komplet skelet såvel som adskillige andre dental-og skeletrester (Whitehite et al. 2009). Ar. ramidus og en ældre, beslægtede arter kendt fra fragmentariske rester, Ar. kadabba (5, 8-5, 2 mya) har reduceret hjørnetænder svarende til Orrorin og Sahelanthropus. Kraniet af Ar. ramidus er temmelig ape-lignende og stort set ligner Sahelanthropus, med en lille chimpanse-størrelse hjerne på 300–350cc (figur 2). Ardipithecus postcranial skelet er spændende., Selv om stærkt opsplittet, bækken gendannede afslører en morfologi er helt forskellig fra den, der lever aber, med en kortere, mere bowl-lignende form, der kraftigt antyder, Ardipithecus gik bipedally; dette er i overensstemmelse med foramen magnum position, hvilket tyder på en opretstående stilling. Imidlertid antyder dens lange forben og fingre og dens divergerende, greb førstetå (hallu.), at Ardipithecus tilbragte meget af sin tid i træerne. Det overordnede indtryk er af en stort set arboreal art, der gik bipedalt, når den turde til jorden.,
Australopithecus
Omkring 4mya vi finde de tidligste medlemmer af slægten Australopithecus, hominins, som var dygtig terrestriske bipeds, men fortsatte med at bruge træer til mad og beskyttelse. De første eksemplarer af Australopithecus blev opdaget i Sydafrika i 1924 (Dart, 1925), og forskningsindsats over de efterfølgende otte årtier har produceret hundredvis af fossiler fra flere arter på lokaliteter i hele det Østlige og Sydlige Afrika., Vi ved nu, at Australopithecus var en meget vellykket slægt, der varede i næsten tre millioner år (Figur 1).
de mest kendte Australopithecus-arter er A. afarensis (3.6–2.9 mya) fra Østafrika og A. Africanus (3.2–2.0 mya) fra Sydafrika., Bækken og nederste del af disse arter, der klart angiver, at de var fuldt ud tobenet: bækkenet er kort og skålformet, at bringe bagdelen muskler rundt til siden af kroppen, som i moderne mennesker, for stammen stabilisering under bipedalism, og den første tå er i overensstemmelse med de andre tæer (Ward, 2002; Harcourt-Smith og Aiello, 2004). Australopithecus-foden kan endda have haft en menneskelignende bue, baseret på analyse af metatarsalerne og de fossiliserede Laetoli-fodspor (Wardard et al. 2011)., Ikke desto mindre, sammenlignet med moderne mennesker, underarmene var lange, og fingre og tæer var lange og noget buede, hvilket antyder, at Australopithecus regelmæssigt brugte træerne til at fodre og måske som et tilflugtssted fra rovdyr om natten. Denne blandede terrestriske & arboreal strategi ville have tjent disse arter godt i de blandede skov-og savannemiljøer, de beboede.
Hjernens størrelse i Australopithecus varierede mellem 390 og 515cc, svarende til chimpanser og gorillaer (Falk et al., 2000), hvilket tyder på, at kognitive evner Stort set lignede levende aber (figur 2). Kropsstørrelsen i Australopithecus var ret lille og seksuelt dimorf, omkring 30 kg for kvinder og 40 kg for mænd (McHenry, 1992). Dette niveau af dimorfisme afspejles ikke i hjørnetænderne, som var små, stump, og monomorf som i tidligere homininer.
i modsætning til hjørnetænderne var molære tænder i Australopithecus meget større end dem fra tidligere homininer og havde tykkere emalje. Dette antyder, at deres kost inkluderede hårde, plantefødevarer af lav kvalitet, der krævede kraftig tygning for at behandle., En undergruppe af Australopithecus, kendt som de “robuste” australopiths (ofte mærket af en separat slægt Paranthropus) på grund af deres enorme tænder og tyggemuskler, tog denne tilpasning til det ekstreme. De fleste Australopithecus-arter blev udryddet af 2 mya, men nogle robuste former varede indtil omkring 1.2 mya i øst og Sydafrika.
slægten Homo
de tidligste fossiler af vores egen slægt, Homo, findes i Østafrika og dateres til 2.3 mya (Kimbel et al. 1997)., Disse tidlige prøver ligner hjerne-og kropsstørrelse som Australopithecus, men viser forskelle i deres molære tænder, hvilket antyder en ændring i kosten. Faktisk ved mindst 1.8 mya brugte tidlige medlemmer af vores slægt primitive stenværktøjer til slagter dyrekroppe og tilføjede energirigt kød og knoglemarv til deres plantebaserede kost.
det ældste medlem af slægten Homo, H. habilis (2.3–1.4 mya) findes i Østafrika og er forbundet med slagtede dyreknogler og enkle stenredskaber (Blumenschine et al. 2003). Dens mere formidable og udbredte efterkommer, H., erectus, findes i hele Afrika og Eurasien og varede fra 1,9 mya til 100 kya, og måske endda senere (Anton, 2003). Ligesom moderne mennesker manglede H. erectus forbentilpasningerne til klatring set i Australopithecus (figur 2). Dens globale ekspansion antyder H. erectus var økologisk fleksibel, med den kognitive kapacitet til at tilpasse sig og trives i meget forskellige miljøer. Ikke overraskende er det med H. erectus, at vi begynder at se en stor stigning i hjernestørrelse, op til 1.250 cc for senere asiatiske prøver (Anton, 2003). Molar størrelse er reduceret i H., erectus i forhold til Australopithecus, hvilket afspejler sin blødere, rigere kost.
Omkring 700 kya, og måske tidligere, H. erectus i Afrika gav anledning til H. heidelbergensis, en art, der meget gerne til os i form af krop proportioner, dental tilpasninger, og de kognitive evner (Rightmire, 2009). H. heidelbergensis, der ofte omtales som en “arkaisk Homo sapiens, var en aktiv big-game hunter, produceret avancerede Levallois stil værktøjer, og mindst 400 kya havde lært at kontrollere ild (Roebroeks og Villa, 2011). Neandertalere (H., neanderthalensis), koldt-tilpasset hominins med stout fysik, komplekse adfærd, og hjerner på størrelse med vores, menes at have udviklet sig fra H. heidelbergensis befolkninger i Europa med mindst 250 kya (Rightmire, 2008; Hublin, 2009).
Fossile og DNA-beviser, der tyder på, vores egen art, H. sapiens, udviklede sig i Afrika 200 kya (Relethford, 2008; Rightmire, 2009), sandsynligvis fra H. heidelbergensis. Den øgede adfærdsmæssige raffinement af H., sapiens, som indikeret af vores store hjerner (1,400 cc) og arkæologiske beviser for et bredere værktøjssæt og kloge jagtteknikker, gjorde det muligt for vores arter at blomstre og vokse på det afrikanske kontinent. Ved 100kya, vores arter spildt i Eurasien, til sidst ekspanderende over hele kloden i Australien og Amerika (DiGiorgio et al. 2009). Undervejs fordrev vores arter andre homininer, de stødte på, herunder neandertalere i Europa og lignende former i Asien. (Bemærk, at ikke alle er enige i denne fortolkning af dataene, se Tryon og Bailey)., Undersøgelser af gammelt DNA ekstraheret fra Neanderthal fossiler antyder, at vores arter lejlighedsvis kan have blandet sig med dem (Green et al., 2010). Vores stigende globale indflydelse fortsætter i dag, da kulturelle innovationer som landbrug og urbanisering former landskabet og arten omkring os.
resum.
udviklingen af vores arter fra en abelignende Miocene forfader var en kompleks proces. Vores afstamning er fuld af sidegrener og evolutionære døde ender, med arter som de robuste australopiths, der varede i over en million år, før de falmede væk., Nogle menneskelige træk, som bipedalisme, udviklede sig meget tidligt, mens andre, som store hjerner, ikke udviklede sig før relativt for nylig. Stadig andre træk, som molstørrelse, udviklede sig i en retning kun for at blive skubbet tilbage senere ved at ændre økologisk pres. I stedet for et kraftigt skib, der kortlægger en lige kurs mod en forudbestemt destination, passer udviklingen af vores afstamning-ja, af enhver Arts afstamning-til billedet af en redningsbåd kastet rundt af skiftende hav af miljøændringer, genetisk held og Geologisk chance. Man undrer sig over, hvor de næste seks millioner år kan tage os.