Obrázek 4.1 (viz rozšířené zobrazení).
z toho vyplývá, že pochopení této synapse vede k pochopení ostatních. Proto budeme nejprve diskutovat o procesu synaptického přenosu na kosterním neuromuskulárním uzlu.,
vlastnosti synaptické křižovatky na neuromuskulární křižovatce jsou zobrazeny na obrázku vlevo. Kosterní svalová vlákna jsou inervovaných motoneurony, jejichž buňky těla jsou umístěna ve ventrální roh míšní. Terminální oblast axonu vede k velmi jemným procesům, které probíhají podél buněk kosterních svalů. Podél těchto procesů jsou specializované struktury známé jako synapse. Konkrétní synapse mezi páteřním motorickým neuronem a buňkou kosterního svalstva se nazývá motorická endplate kvůli své specifické struktuře.,
synapse na neuromuskulární křižovatce má tři charakteristické rysy chemických synapsí v nervovém systému. Za prvé, existuje zřetelné oddělení mezi presynaptickou a postsynaptickou membránou. Prostor mezi nimi je známý jako synaptická štěrbina. Prostor nám říká, musí existovat nějaký zprostředkující signální mechanismus mezi presynaptického neuronu a postsynaptického neuronu aby měl tok informací přes synaptické štěrbiny. Za druhé, existuje charakteristická vysoká hustota malých sférických váčků., Tyto synaptické váčky obsahují neurotransmiterové látky. Synapse jsou také spojeny s vysokou hustotou mitochondrií. Za třetí, ve většině případů, tam je charakteristické ztluštění postsynaptické membráně, což je dáno přinejmenším zčásti tím, že postsynaptické membrány má vysokou hustotu specializované receptory, které se vážou chemické vysílač látky uvolňované z presynaptického neuronu. Další podrobnosti o morfologických rysech synaptických křižovatek jsou uvedeny v kapitole 8 A kapitole 10.
4.,2 Fyziologie Synaptického Přenosu na Nervosvalové ploténce
Obrázek 4.2
na obrázku vpravo ukazuje ve velmi schematický způsob, jak je možné studovat fyziologii synaptického přenosu na nervosvalové ploténce kosterních ve velkém detailu. Kus svalu a jeho připojený nerv jsou umístěny v malé experimentální komoře naplněné vhodným roztokem vyzvánění. Klidový potenciál svalové buňky je zaznamenán mikroelektrodou. Elektrody jsou také umístěny na povrchu nervového axonu., Krátké elektrické šoky způsobují iniciaci akčních potenciálů, které se šíří do synaptického terminálu.
níže uvedený obrázek ilustruje dva typy potenciálních změn, které byly zaznamenány v takovém izolovaném nervově-svalovém přípravku. Experiment také ukazuje vlastnosti silný lék, kurare, který se ukázal být velmi užitečné při studiu procesu synaptického přenosu na nervosvalové ploténce kosterních. Část a ilustruje posloupnost potenciálních změn zaznamenaných ve svalové buňce v důsledku stimulace motorického axonu., Šipka označuje bod v čase, kdy je šok přiveden do motoru axon. Všimněte si, že po šoku je klidová doba. Zpoždění je způsobeno časem potřebným k tomu, aby se akční potenciál v motoru axon šířil z místa jeho zahájení. Po zpoždění jsou ve svalové buňce zaznamenány dva typy potenciálů. Za prvé, existuje relativně pomalu se měnící potenciál, který bude předmětem následující diskuse., Pokud je tento pomalý počáteční potenciál dostatečně velký, protože je obvykle v buňkách kosterních svalů, je ve svalové buňce vyvolán druhý potenciál, akční potenciál.
4.3
Akční potenciály v kosterní svalové buňky jsou v důsledku iontové mechanismy podobné těm, diskutovali dříve. Konkrétně dochází ke změně propustnosti na+ závislé na napětí a následnému zpožděnému zvýšení propustnosti k+. (Pro buňky hladkého svalstva a buňky srdečního svalu jsou iontové mechanismy odlišné.,)
základní událost, která spouští akční potenciál může být odhalena s využitím kurare, šípový jed používají některé jihoamerických Indiánů. Nízká dávka přípravku curare (část B) snižuje základní událost, ale stále není dostatečně snížena, aby klesla pod prahovou hodnotu. Pokud je podávána poněkud vyšší dávka přípravku curare (část C), pomalá podkladová událost se stává subthreshold. Základní signál je známý jako potenciál desky (EPP), protože to je potenciální změna zaznamenána u motoru, zavazadlového prostoru. Obecně se nazývá excitační postsynaptický potenciál (EPSP).,
Curare blokuje potenciál endplate, protože je konkurenčním inhibitorem acetylcholinu (ACh), vysílače uvolněného na presynaptickém terminálu. Curare neblokuje napěťově závislou vodivost Na+ nebo vodivost závislou na napětí k+, která je základem potenciálu působení svalů. Curare ovlivňuje stimul (EPSP), který obvykle vede k zahájení svalového akčního potenciálu. Zvíře, které je otráveno curare, se udusí, protože proces neuromuskulárního přenosu v respiračních svalech je blokován.,
obvykle je velikost potenciálu koncové desky poměrně velká. Amplituda potenciálu koncové desky je asi 50 mV, ale k dosažení prahu je zapotřebí pouze asi 30 mV. Dalších 20 mV se nazývá bezpečnostní faktor. Proto, i když čelní desky potenciál, aby se stal poněkud menší (např. 40 mV amplituda) z důvodu únavy, EPP dosáhne práh a one-to-one vztah mezi akčním potenciálem v motoru axon a akční potenciál ve svalové buňce by být zachována.
obrázek 4.4
4.,3 propagace EPP
jaké jsou vlastnosti EPP a jak se porovnává s vlastnostmi akčního potenciálu?
je potenciál endplate způsoben změnou na+ a k+ permeabilities závislou na napětí, jako je akční potenciál?
je potenciál endplate propagován všemi nebo ničím podobným akčnímu potenciálu?
obrázek vlevo ilustruje experiment, který zkoumá šíření potenciálu endplate. Svalové vlákno je opakovaně nabodnuto elektrodami v intervalech 1 mm., (Všimněte si, že čelní desky potenciál je malý, protože tento experiment se provádí v přítomnosti nízké koncentrace kurare, takže čelní desky potenciál může být zaznamenán bez komplikací spouští akční potenciál.) Potenciál endplate se nepropaguje celoplošným způsobem. Šíří se podél svalu, ale dělá to s poklesem., To znamená, že se šíří z čelní desky potenciál ze svého místa zasvěcení do dalších míst podél svalových buněk dochází pasivně a s decrement, jen jako udržovací funkce možné změny v jedné části axonu se šíří podél axonu, nebo jen jako změna teploty na jeden bod na kovové tyči se šíří podél tyče.
4.4 Přehled o Sledu Událostí Hlubších EPP
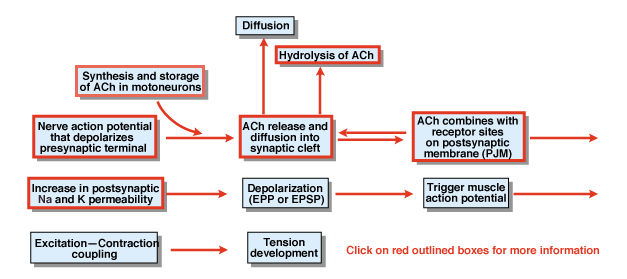
Obrázek 4.5
Jaké jsou další kroky v procesu chemické synaptické transmise? Obrázek 4.,5 poskytuje přehled. Nervový akční potenciál, který je iniciován v buněčném těle páteřního motorického neuronu, šíří ventrální kořeny a nakonec napadá synaptické terminály motorických neuronů. V důsledku akčního potenciálu se chemický vysílač acetylcholin (ACh) uvolňuje do synaptické štěrbiny. ACh difunduje přes synaptickou štěrbinu a váže se na speciální receptory na postsynaptické nebo postjunktivní membráně. Vazba ACh na jeho receptory vytváří konformační změnu v membránovém kanálu, který je specificky propustný jak pro Na+, tak pro k+., V důsledku zvýšení propustnosti Na+ a k+ dochází k depolarizaci postsynaptické membrány. Tato depolarizace se nazývá potenciál endplate nebo obecněji EPSP. Pokud je EPSP dostatečně velký, protože je obvykle na neuromuskulární křižovatce, vede k zahájení akčního potenciálu ve svalové buňce. Akční potenciál iniciuje proces excitační kontrakční vazby a vývoj napětí. Doba trvání potenciálu endplate je asi 10 msec.
dva faktory řídí trvání EPSP na neuromuskulární křižovatce., Za prvé, ACh je odstraněn difúzí. Za druhé, látka v synaptické štěrbině, nazývaná acetylcholinesteráza (AChE), hydrolyzuje nebo rozkládá ACh. AChE je jedním z nejúčinnějších enzymů známých. Jedna molekula AChE může hydrolyzovat 600 000 molekul ACh za minutu.
4.5 Úloha AChE
Obrázek 4.6
důležitou skupinu látek, z nichž jedna je neostigmin, inhibuje účinek AChE. Neostigmin blokuje působení bolesti, a tím činí potenciál endplate větší a delší dobu trvání., Tento obrázek znázorňuje dva endplate potenciály. Jeden byl zaznamenán ve fyziologickém roztoku a curare a druhý zaznamenán po přidání neostigminu do roztoku. (Curare je přidán tak, aby vlastnosti EPP mohly být studovány bez vyvolání akčního potenciálu ve svalové buňce.) Po aplikaci neostigminu je potenciál endplate mnohem větší a delší.
4.6 Myasthenia Gravis
Myasthenia gravis je spojena s těžkou svalovou slabost, protože snížením počtu acetylcholinových receptorů ve svalové buňce., Pokud je potenciál koncové desky menší, potenciál koncové desky nedosáhne prahové hodnoty. Pokud nedosáhne prahu, nebude ve svalové buňce žádný akční potenciál a žádná kontrakce svalu, která způsobuje svalovou slabost. Neostigmin a další inhibitory AChE se používají k léčbě pacientů s myasthenia gravis. Tato činidla činí množství acetylcholinu, které se uvolňuje účinněji, dosáhne zbývajících acetylcholinových receptorů.
4.,6b nervová činidla
přestože inhibitory AChE mají důležitou terapeutickou hodnotu, některé inhibitory byly a stále se používají jako jedy. Některé inhibitory bolesti, jako je Soman a Sarin, tvoří poměrně nevratný blok bolesti. Tento blok vede k extrémním úrovním ACh v synaptické štěrbině. Jedinci tak otrávení umírají na záchvaty a svalovou spasticitu včetně respiračních svalů.
obrázek 4.7
4.,7 Iontoforéza ACh,
Iontoforéza je zajímavá technika, která může být použita k dalšímu testování hypotézy, že ACh je neurotransmiter, látka na nervosvalové ploténce. Jestli ACh je vysílač, který je propuštěn tento synapse, dalo by se předpovídat, že by mělo být možné nahradit umělou aplikace vysílač pro normální uvolnění vysílače. Protože ACh je pozitivně nabitá molekula, může být vytlačena z mikroelektrody, aby simulovala uvolňování ACh z presynaptického terminálu.
obrázek 4.,8
do blízkosti neuromuskulárního spojení lze skutečně aplikovat minutové množství ACh. Obrázek 4.8 porovnává EPP produkovaný stimulací motorového axonu a reakcí na ejekce ACh. Potenciální změna vypadá téměř identicky s potenciálem endplate produkovaným normálním uvolněním ACh. Tento experiment poskytuje experimentální podporu konceptu, že ACh je přirozený vysílač v této synapse.,
reakce na vyhození ACh má některé další zajímavé vlastnosti, které jsou všechny v souladu s cholinergní charakter synapse na kosterní nervosvalové ploténce. Neostigmin činí odpověď na iontoforézu ACh delší a větší. Curare snižuje odezvu, protože soutěží s normální vazbou ACh. Pokud je ACh vysunut do svalové buňky, nic se nestane, protože receptory pro acetylcholin nejsou uvnitř; jsou na vnější straně svalové buňky., Aplikace acetylcholinu do regionů svalu od konce-deska produkuje žádnou odpověď, protože receptory pro ACh jsou soustředěny v synaptické oblasti.
Chcete-li otestovat své porozumění, zvažte, jak by agent, jako je TTX, ovlivnil tvorbu EPP i reakci svalového vlákna na iontoforetickou aplikaci ACh? TTX nemá žádný vliv na odpověď na ACh, ale blokuje EPP. Důvod, proč reakce na ACh není ovlivněna, je jasný, ale mnozí očekávají, že pokud zde nebude žádný účinek, neměl by mít žádný vliv ani na EPP., Tetrodotoxin neovlivňuje vazbu acetylcholinu na receptory, a proto neovlivní odpověď na přímou aplikaci ACh. Tetrodotoxin však ovlivní schopnost akčního potenciálu vyvolaného v motorickém axonu. Pokud nelze v motoru axon vyvolat akční potenciál, nemůže to způsobit uvolnění vysílače. Tetrodotoxin by tak EPP zcela zrušil. Blok by nebyl způsoben blokem ACh receptorů, ale spíše blokem nějakého kroku před uvolněním vysílače.
4.,8 iontové mechanismy EPP
Bernard Katz a jeho kolegové byli průkopníky ve vyšetřování mechanismů synaptického přenosu na neuromuskulární křižovatce. Navrhli, že kanál otevřený ACh byl ten, který měl stejnou propustnost jak Na+, tak k+. Protože to bylo stejně propustné pro na+ a k+, Katz navrhl, že v důsledku otevření těchto kanálů by se membránový potenciál pohyboval směrem k 0 mV. (Hodnota alfa v GHK rovnici rovnající se jedné, která při substituci do rovnice poskytuje potenciál asi 0 mV.,)
obrázek 4.9
experiment zobrazený na obrázku vlevo testuje tento koncept. Svalová buňka byla proniknuta nahrávací elektrodou a další elektrodou, která může být připojena k vhodnému zdroji potenciálu, aby se uměle změnil membránový potenciál. Normálně je membránový potenciál opět asi -80 mV, přidává se malé množství curare, takže EPP je malý. Katz si v těchto experimentech všiml, že velikost EPP se dramaticky změnila v závislosti na potenciálu svalové buňky., Pokud je membránový potenciál přesunut na 0 mV, není zaznamenána žádná potenciální změna. Pokud je membránový potenciál + 30 mV, EPP je obrácený. Takže tři různé podněty produkují endplate potenciály, které se od sebe velmi liší.
nedostatek odpovědi, pokud je potenciál na 0 mV, je zvláště informativní. Zvažte, proč není zaznamenána žádná potenciální změna. Předpokládá se, že vysílač je uvolňován a vázán na receptory., Jednoduché vysvětlení nedostatku potenciální změny spočívá v tom, že potenciál, na který se otevření ACh kanálů snaží dosáhnout, již byl dosažen. Pokud je membránový potenciál pozitivnější než 0 mV,pak je EPP obrácen. Bez ohledu na to, jaký je potenciál, změna propustnosti má tendenci pohybovat membránovým potenciálem směrem k 0 mV! Pokud je klidový potenciál zápornější než 0 mV, dochází k vychýlení nahoru. Pokud je pozitivnější, dochází k vychýlení směrem dolů. Pokud je již na 0 mV, nedochází k vychýlení.
obrázek 4.,10
tento potenciál se také nazývá potenciál obrácení, protože se jedná o potenciál, při kterém se změní znaménko synaptického potenciálu. Experiment naznačuje, že v důsledku ACh vazby na receptory se specifické kanály stávají stejně propustnými pro Na+ a k+. Tato změna propustnosti má tendenci posouvat membránový potenciál odkudkoli je zpočátku směrem k novému potenciálu 0 mV.
proč normální potenciál endplate nikdy nedosáhne 0 mV?, Jedním z důvodů je, že sled změn propustnosti, které jsou základem akčního potenciálu,“ zaplavuje “ změny produkované EPP. Ale i kdyby nebyl spuštěn akční potenciál, EPP by stále nedosáhl 0 mV. Je to proto, že ACh kanály jsou jen malou část celkového počtu kanálů ve svalových vláknech. K + kanály, které poskytují svalovou buňku svým klidovým potenciálem, jsou také přítomny. Jejich úkolem je pokusit se udržet buňku v klidovém potenciálu.,
kanál otevřel ACh je členem obecné třídy kanály, tzv. ligand-gated kanály nebo ionotropní receptory. Jak je znázorněno na obrázku 4.10, místo vazby vysílače je součástí samotného kanálu. Jako výsledek vysílač vazba na receptor (obvykle dvě molekuly jsou nezbytné), je konformační změna proteinu, což umožňuje pórů regionu otevřít a ionty proudit dolů jejich elektrochemické gradienty. Další podrobnosti o kanálu jsou uvedeny v kapitole 11.,
Otestujte si Své Znalosti
- na Otázku 1
- B
- C
- D
Na desky potenciál v kosterní svalové buňky by v zásadě mohlo být produkován snížená propustnost pro které z následujících iontů(y)? (Předpokládejme, že existuje konečná počáteční propustnost pro každý z níže uvedených iontů a že jsou přítomny fyziologické gradienty koncentrace.):
a. na +
b. Na+ a Ca2+
C. Ca2 +
d., K +
potenciál endplate v buňce kosterního svalstva by mohl být v zásadě produkován sníženou propustností pro který z následujících iontů(iontů)? (Předpokládejme, že existuje konečná počáteční propustnost pro každý z níže uvedených iontů a že jsou přítomny fyziologické gradienty koncentrace.):
a. Na+ tato odpověď je nesprávná.
potenciál koncové desky je depolarizace, která je normálně produkována současným zvýšením propustnosti pro ionty sodíku a draslíku., Pokud by došlo k selektivnímu snížení propustnosti sodíku, takové snížení by nevedlo k depolarizaci. Spíše by to ve skutečnosti vedlo k hyperpolarizaci, protože alfa v GHK rovnici by byla snížena. Poměr propustnosti pro sodné a draselné by být upřednostňován na draslík propustnost, pohybující se membránový potenciál směrem k draslíku rovnovážný potenciál a výrobu hyperpolarizace.
b. Na + a Ca2+
C. Ca2 +
d., K +
potenciál endplate v buňce kosterního svalstva by mohl být v zásadě produkován sníženou propustností pro který z následujících iontů(iontů)? (Předpokládejme, že existuje konečná počáteční propustnost pro každý z níže uvedených iontů a že jsou přítomny fyziologické gradienty koncentrace.):
a. na +
b. Na+ a Ca2 + tato odpověď je nesprávná.
Pokud propustnost pro sodík a vápník byly sníženy jejich důsledky budou podobné, že v možnost a. Snížení propustnosti sodný, sám by spíše hyperpolarizace buňky., Podobně snížení propustnosti pro vápník může také hyperpolarizovat buňku. Rovnovážný potenciál vápníku je velmi pozitivní hodnota, a pokud by došlo k nějaké tonické klidové propustnosti pro vápník, tato propustnost by přispěla k tonické depolarizaci membránového potenciálu. Snížení propustnosti vápníku by tedy odstranilo tento tonický depolarizační účinek a vedlo by k hyperpolarizaci.
C. Ca2 +
d., K +
potenciál endplate v buňce kosterního svalstva by mohl být v zásadě produkován sníženou propustností pro který z následujících iontů(iontů)? (Předpokládejme, že existuje konečná počáteční propustnost pro každý z níže uvedených iontů a že jsou přítomny fyziologické gradienty koncentrace.):
a. na +
b. Na+ a Ca2+
C. Ca2 + tato odpověď je nesprávná.
snížení propustnosti vápníku by samo o sobě vedlo k hyperpolarizaci. Viz logika odpovědi na volbu b.
d., K +
potenciál endplate v buňce kosterního svalstva by mohl být v zásadě produkován sníženou propustností pro který z následujících iontů(iontů)? (Předpokládejme, že existuje konečná počáteční propustnost pro každý z níže uvedených iontů a že jsou přítomny fyziologické gradienty koncentrace.):
a. na +
B. na+ a Ca2+
C. Ca2 +
D. K + tato odpověď je správná!
snížení propustnosti draslíku by vedlo k depolarizaci podobné potenciálu koncové desky., Je tomu tak proto, že v klidu existuje tonická propustnost pro draslík a sodík. Vysoká propustnost pro draslík má tendenci udržovat membránový potenciál v blízkosti rovnovážného potenciálu draslíku. Případě, že klidová propustnosti se snížil, alfa v Goldman rovnice by se stal větší hodnotu, pohybující se membránový potenciál trochu blíže k sodíku rovnovážný potenciál (depolarizace).