Úvod
Darwin je skvělý pohled, a jednotícím principem v biologii dnes, je, že všechny druhy jsou navzájem související, jako sestry, bratranci a vzdálení příbuzní v obrovské rodinného stromu života. Důsledky jsou dechberoucí; kdybychom mohli cestovat zpět dostatečně daleko v čase, našli bychom společné předky mezi sebou a každým jiným živým organismem, od dikobrazů po plameňáky až po Kaktusy., Naše bezprostřední evoluční rodina se skládá z hominoids, skupina primátů, která zahrnuje „menší opic“ (siamangs a gibbons), stejně jako „lidoopy“ (šimpanzi, bonobové, gorily a orangutani). Mezi velkými lidoopy jsou našimi nejbližšími příbuznými šimpanzi a bonobové (Obrázek 1). Fosilní záznam, spolu s studie lidské a opičí DNA, naznačují, že lidé sdílí společného předka se šimpanzi a bonobové někdy kolem 6 miliony let (mya)., Začneme diskusi o náš vývoj druhů v Africe, téměř na konci geologického období, známého jako Miocénu, těsně předtím, než naše linie rozešly od šimpanzi a bonobové.
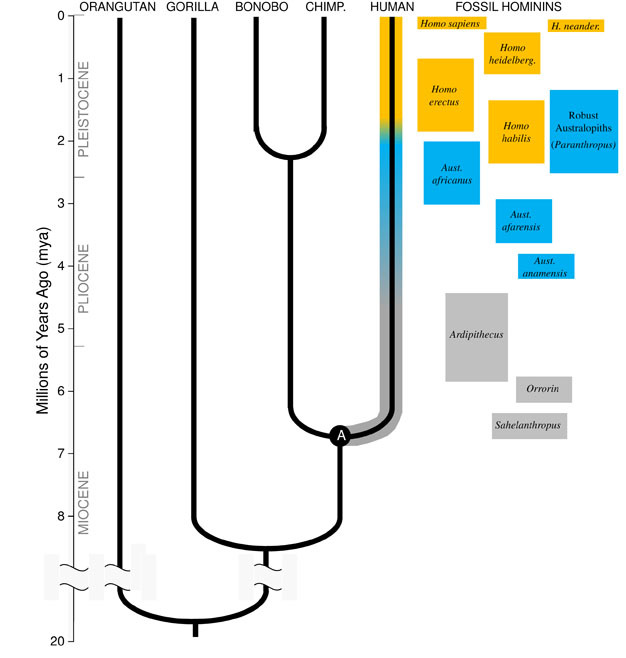
Miocénu Původ Hominin Linie
aby bylo možné pochopit vývoj všech druhů, musíme nejprve vytvořit jeho předků státu: jaké zvíře se to vyvíjet? Pro naši linii to vyžaduje, abychom se pokusili rekonstruovat posledního společného předka lidí a šimpanzů (označeného „A“ na obrázku 1). Lidský šimpanz poslední společný předek (HC-LCA) je druh, od kterého se rod homininů a šimpanz & bonobo lineage rozcházel., Hominins jsou druhy na naší pobočce hominoidní strom po rozdělení s šimpanzem & bonobo linka, včetně všech vyhynulých druhů a evoluční boční větve (viz Obrázek 1).
v Miocénu byla velká rozmanitost druhů opic s desítkami druhů známých z fosilních záznamů po celé Africe, Evropě a Asii. Tyto druhy se lišily svou anatomií a ekologií a není jasné, který z dosud objevených fosilních druhů představuje HC-LCA (Kunimatsu et al. 2007; Young and MacLatchy, 2004)., Nicméně, z fosilních a srovnávacích důkazů víme, že to bylo mnohem více podobné živým lidoopům než živým lidem. HC-LCA by měl opičí mozek a tělo, s relativně dlouhými pažemi a prsty a uchopovací nohou, která mu umožnila pícninu ve stromech. Psí zuby byly pravděpodobně velké a ostré, jak je vidět u několika miocénních hominoidů. Navíc, špičáky byly pravděpodobně sexuálně dimorfní, s muži mají mnohem větší špičáky, než ženy, jak je vidět mezi živými lidoopy a Miocénu zkameněliny., Jako žijící opice by šel quadrupedally (na všech čtyřech), když na zemi, a jeho strava by se skládala téměř výhradně z rostlinných potravin, zejména ovoce a listí.
časné homininy
změny z anatomie podobné lidoopům jsou rozeznatelné u hominoidních fosilií z pozdního miocénu v Africe. Některé hominoid druhů z tohoto období vykazují rysy, které jsou typické pro člověka, ale nejsou vidět v jiných živých opic, což vede paleoantropologové odvodit, že tyto fosilie představují prvních členů hominin linie., První lidské rysy, které se objevují ve fosilním záznamu homininu, jsou bipedální chůze a menší, tupé špičáky.
nejstarší homininy, které jsou v současné době známy, jsou Sahelanthropus tchadensis z Čadu (Brunet et al. 2005) a Orrorin tugenensis z Keni (Senut et al. 2001). Sahelanthropus, datován mezi 6 a 7 mya, je známý z převážně úplné lebky a některých dalších fragmentárních pozůstatků. Jeho velikost mozku, 360cc, je v rozmezí vidět ve šimpanzi, a lebka, masivní nadočnicové, podobné v tloušťce samec gorily (Brunet et al. 2005)., Nicméně, pozice a orientace foramen magnum, otvor v základně lebky, jehož prostřednictvím míchy prochází, naznačuje, že Sahelanthropus se postavil a šel bipedally, s jeho páteře držen svisle, jako u moderních lidí, spíše než vodorovně jako u opic a jiných čtvernožců (Zollikofer et al. 2005). Orrorin je znám především z postkraniálních fosilií, včetně částečné stehenní kosti. Proximální část stehenní kosti vykazuje podobnosti s podobnostmi moderních lidí, což naznačuje, že druh byl bipedální (Pickford et al. 2002)., Nebyly nalezeny žádné lebky Orrorinu, a proto je jeho kraniální morfologie a velikost mozku nejistá. V obou Orrorin a Sahelanthropus psí zuby samci jsou větší a špičatější než u moderních lidí, ale jsou malé a tupé oproti špičáky samců opic. To naznačuje, že psí pohlavní dimorfismus — a potažmo hospodářské soutěže mezi samci o přístup k páření samic — byla snížena v těchto raných hominins ve srovnání s lidoopy.
zdaleka nejznámějším časným homininem je Ardipithecus ramidus, a 4.,4 miliony let starý druh z Etiopie, který je známý z téměř kompletní kostry, stejně jako mnoho dalších zubních a kosterních pozůstatků (White et al. 2009). Ar. ramidus a starší, příbuzné druhy známé z fragmentárních pozůstatků, Ar. kadabba (5,8-5,2 mya), mají snížené špičáky podobné špičkám Orrorinu a Sahelanthropu. Lebka Ar. ramidus je spíše opičí a široce podobný mozku Sahelanthropus, s malým mozkem velikosti šimpanze 300–350cc (Obrázek 2). Ardipithecus postkraniální kostra je zajímavá., I když moc roztříštěné, pánve zotavil odhaluje morfologie naprosto odlišná od živých opic, s kratší, více miska-jako tvar, který silně naznačuje, Ardipithecus šel bipedally; to je v souladu s foramen magnum pozici, což naznačuje, vzpřímené držení těla. Jeho dlouhé přední končetiny a prsty a divergentní uchopení prvního prstu (hallux) však naznačují, že Ardipithecus strávil většinu času ve stromech. Celkový dojem je z převážně arboreálního druhu, který šel bipedálně, kdykoli se odvážil k zemi.,
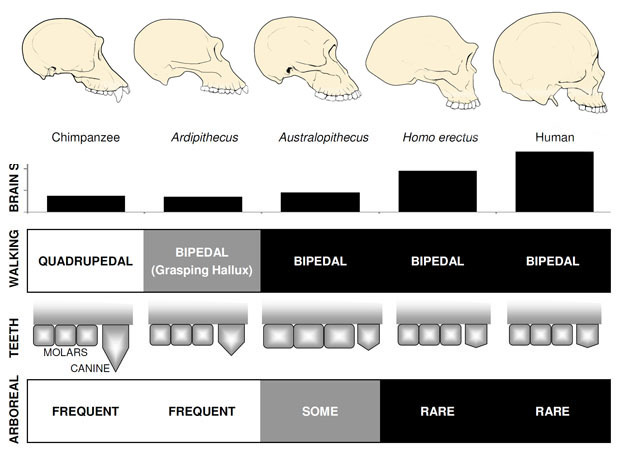
Australopithecus
Kolem 4mya najdeme nejstarší členové rodu Australopithecus, hominins, které byly adept pozemní dvounožci, ale i nadále používat stromy za jídlo a ochranu. První exempláře Australopithecus byl objeven v Jižní Africe v roce 1924 (Dart 1925), a úsilí v oblasti výzkumu, během následujících osm desetiletí vyrábí stovky zkamenělin z několika druhů na místech po celé Východní a Jižní Africe., Nyní víme, že Australopithecus byl velmi úspěšný rod, který přetrvával téměř tři miliony let (Obrázek 1).
nejznámějším druhem Australopithecus je a. afarensis (3.6–2.9 mya) z východní Afriky a a.africanus (3.2–2.0 mya) z Jižní Afriky., Pánev a dolní končetiny těchto druhů jasně ukazuje, že oni byli plně bipedální: pánev je krátké a ve tvaru mísy, čímž hýžďové svaly po straně těla, jako u moderních lidí, pro kufr stabilizace během bipedalism, a první prst je v souladu s ostatními prsty (Ward, 2002; Harcourt-Smith a Aiello, 2004). Noha Australopithecus mohla mít dokonce lidský oblouk založený na analýze metatarzálů a zkamenělých laetoliových stop (Ward et al. 2011)., Nicméně, ve srovnání s moderními lidmi, předloktí byly dlouhé a prsty a prsty byly dlouhé a poněkud zakřivené, což naznačuje, že Australopithecus pravidelně používají stromy píce a možná i jako úkryt před predátory v noci. Tato smíšená terestrická strategie& by těmto druhům dobře sloužila ve smíšeném prostředí lesů a savan, které obývali.
velikost mozku u Australopithecus se pohybovala mezi 390 a 515 ccm, podobně jako šimpanzi a gorily (Falk et al., 2000), Což naznačuje, že kognitivní schopnosti byly široce podobné živým lidoopům (Obrázek 2). Velikost těla v Australopithecus byla poměrně malá a sexuálně dimorfní, asi 30 kg pro ženy a 40 kg pro muže (McHenry, 1992). Tato úroveň dimorfismu se neodráží v špičácích, které byly malé, tupé a monomorfní jako u dřívějších homininů.
Na rozdíl od špičáků byly molární zuby v Australopithecusu mnohem větší než zuby dřívějších homininů a měly silnější smalt. To naznačuje, že jejich strava zahrnovala tvrdé, nekvalitní rostlinné potraviny, které vyžadovaly silné žvýkání ke zpracování., Podskupina Australopithecus, známá jako“ robustní “ australopiths (často označená samostatným rodem Paranthropus) kvůli jejich obrovským zubům a žvýkacím svalům, vzala tuto adaptaci do extrému. Většina druhů Australopithecus byla vyhubena 2 mya, ale některé robustní formy přetrvávaly až do asi 1.2 mya ve východní a Jižní Africe.
Rodu Homo
nejstarší zkameněliny z našeho vlastního rodu Homo, se nacházejí ve Východní Africe a ze dne na 2,3 mya (Kimbel et al. 1997)., Tyto rané vzorky jsou podobné velikosti mozku a těla jako Australopithecus, ale vykazují rozdíly v jejich molárních zubech, což naznačuje změnu stravy. Skutečně, alespoň 1,8 miliony let, první členové našeho rodu byly pomocí primitivní kamenné nástroje, aby řezník zvířecích těl, přidání energie bohaté na maso a kostní dřeně, aby jejich rostlinnou stravu.
nejstarší člen rodu Homo, H. habilis (2.3–1.4 mya) se nachází ve Východní Africe a je spojena s zmasakrovali zvířecí kosti a jednoduché kamenné nástroje (Blumenschine et al. 2003). Jeho impozantnější a rozšířenější potomek h., erectus se nachází v celé Africe a Eurasii a přetrvával od 1,9 mya do 100 kya a možná i později (Anton, 2003). Stejně jako moderní lidé, h.erectus postrádal přední končetiny adaptace pro lezení vidět v Australopithecus (Obrázek 2). Jeho globální expanze, naznačuje, H. erectus byl ekologicky flexibilní, s kognitivní schopnost přizpůsobit se a prosperovat v diametrálně odlišné prostředí. Není divu, že S H. erectusem začínáme vidět velký nárůst velikosti mozku, až 1 250 ccm pro pozdější asijské exempláře (Anton, 2003). Molární velikost je snížena v H., erectus vzhledem k Australopithecus, odrážející jeho měkčí, bohatší stravu.
Kolem 700 kya, a možná i dříve, H. erectus v Africe vedla k H. heidelbergensis, druh, velmi podobně jako nás, pokud jde o tělesné proporce, zubní úpravy, a kognitivní schopnosti (Rightmire, 2009). H. heidelbergensis, často odkazoval se na jako „archaické“ Homo sapiens, byl aktivní a big-game lovcem, vyrábí sofistikované Levallois styl, nástroje a nejméně 400 kya se naučil ovládat oheň (Roebroeks a Vily, 2011). Neandrtálci (H., neanderthalensis), zima-přizpůsobit hominins s korpulentní postavy, složité chování, a mozky podobné velikosti, aby nás, jsou myšlenka k se vyvinuli z H. heidelbergensis populace v Evropě nejméně 250 kya (Rightmire, 2008; Hublin, 2009).
Fosilní a DNA důkazy naznačují, našeho druhu H. sapiens, vyvinul v Africe 200 kya (Relethford, 2008; Rightmire, 2009), pravděpodobně z H. heidelbergensis. Zvýšená behaviorální sofistikovanost h., sapiens, jak naznačují naše velké mozky (1 400 ccm) a archeologické důkazy o širší sadě nástrojů a chytrých loveckých technikách, umožnily našemu druhu vzkvétat a růst na africkém kontinentu. Do 100kya se náš druh rozlil do Eurasie a nakonec se rozšířil po celém světě do Austrálie a Ameriky (DiGiorgio et al. 2009). Cestou náš druh vysídlil další homininy, se kterými se setkali, včetně neandrtálců v Evropě a podobných forem v Asii. (Všimněte si, že ne všichni souhlasí s touto interpretací údajů, viz Tryon a Bailey)., Studie starověké DNA extrahované z neandrtálských fosilií naznačují, že se s nimi náš druh mohl občas spojit (Green et al., 2010). Náš rostoucí globální dopad pokračuje i dnes, protože kulturní inovace, jako je zemědělství a urbanizace, formují krajinu a druhy kolem nás.
souhrn
vývoj našeho druhu z předka miocénu podobného opicím byl složitý proces. Naše linie je plná postranních větví a evolučních slepých uliček, s druhy, jako jsou robustní australopiths, které přetrvávaly více než milion let, než zmizely., Některé lidské rysy, jako je bipedalismus, se vyvinuly velmi brzy, zatímco jiné, stejně jako velké mozky, se vyvíjely až relativně nedávno. Ještě další rysy, jako je molární velikost, se vyvinuly v jednom směru, aby byly později zatlačeny změnou ekologických tlaků. Spíše než silný loď grafů přímý směr k nějaký předem stanovený cíl, evoluce našeho rodu — vlastně všech druhů rodu — odpovídá obraz záchranného člunu hodil o přesun moří změn životního prostředí, genetické štěstí, a geologické šanci. Člověk se diví, kam nás dalších šest milionů let může vzít.